

Introduction
The cultivated peanut (Arachis hypogaea L.) is a major crop in most tropical and subtropical areas of the world and provides a significant source of oil and protein to large segments of the populations of Asia, Africa and South America. In the U. S., peanut is a high-value cash crop of regional importance, with major production areas concentrated in the Southeast.
The genus Arachis is native to South America, with central Brazil postulated as the center of origin. The cultivated peanut and one other allotetraploid species (Arachis monticola Krapov. et Rig.) have been assigned to section Arachis along with at least 25 diploid species (Krapovickas and Gregory, 1994). Although large germplasm collections of both the cultivated and related wild species are available, peanut breeders have traditionally relied on crossing elite by elite germplasm for developing improved cultivars (Holbrook and Stalker, 2003). As a result, the germplasm base of peanut cultivars is extremely narrow and low levels of genetic variability at the molecular level have been shown by isozyme, Restriction Fragment Length Polymorphism (RFLP), Randomly Amplified Polymorphic DNA (RAPD), and Single Sequence Repeat (SSR) analyses (Patterson et al., 2004). Conversely, abundant polymorphisms have been detected among related wild species in section Arachis with these same marker types (Stalker and Mozingo, 2001), representing a valuable source of desirable traits not available in the cultivated gene pool.
Molecular markers offer plant geneticists and breeders a set of genetic tools that are abundant, non-deleterious and reliable. Marker systems have been successfully used over the last several decades to construct genetic maps, assess genetic diversity, and locate genes of interest. The advent of the RAPD assay (Williams et al., 1990) provided an efficient method to detect DNA polymorphisms and generate a large number of molecular markers for genomic applications. RAPD markers for mapping and analysis of genetic diversity have been reported for a wide variety of plants including tobacco (Nicotiana spp.) (Lin et al., 2001), plantain (Musa spp.) (Ude et al., 2003), potato (Solanum tuberosum L.) (Sun et al., 2003) and barley (Hordeum vulgare) (Fernandez et al., 2002).
Genetic maps are normally constructed from a single segregating population, but it is expected that some of the information can be applied to other crosses or species. In peanut, two low density genetic maps have been published by utilizing variation in Arachis species, including one from diploids (Halward et al., 1993) and the second from tetraploid crosses with A. hypogaea (Burow et al., 2001). In the case of polyploid species such as peanut, it is often more efficient to construct initial maps in diploid relatives. Since the genomes of diploid and closely related polyploid species should be similar, one can predict the positions of loci in polyploids based on diploid maps and then search for markers associated with single gene or quantitative trait loci (QTLs) with DNA clones that are most likely to be spaced at regular intervals throughout the polyploid's genome. This strategy can save considerable time and resources versus making de novo maps in polyploids. The approach has been employed in wheat (Guyomarc'h et al., 2002; Gill et al., 1991), alfalfa (Echt et al., 1993; Brummer et al., 1993), and oat (Yu and Wise, 2000).
Halward et al. (1993) constructed an RFLP map in peanut from an F2 population derived from an interspecific cross between the two diploid species (A. stenosperma Krapov. and W.C. Gregory and A. cardenasii Krapov. and W.C. Gregory). Due to the dominant nature of RAPD markers and, to take advantage of the speed with which large numbers of polymorphic markers can be identified, a RAPD map utilizing a backcross population derived from the same interspecific cross [A. stenosperma × (A. stenosperma × A. cardenasii)] was constructed. By establishing linkage relationships with common RFLP markers, or bridge markers, RAPD markers could then be positioned on the peanut core RFLP map. A comparison with the previous map (Halward et al., 1993) is based on a common set of probes and RAPD primers mapped in both studies.
Materials and Methods
Plant Material
A backcross population of 44 plants derived from the cross [A. stenosperma × (A. stenosperma × A. cardenasii)] represented the mapping population (referred to as the BC population). A single F1 plant was used as a male parent and backcross seeds were obtained by hand emasculation and controlled pollination. Plants were grown and maintained in greenhouse facilities at North Carolina State Univ., Raleigh, NC.
RFLP Analyses
Forty-one RFLP probes of known map position (Halward et al., 1993) were selected to provide coverage of all linkage groups and used to screen the BC population. They also provided anchor points to compare the BC and F2 maps. DNA isolation, digestion, Southern blotting and hybridization procedures were completed as described by Kochert et al. (1991).
RAPD Analyses
DNA isolation, reaction mixtures, PCR amplification conditions and DNA size fractionation were performed as described by Garcia et al. (1995). To construct a RAPD map, a total of 428 10-base random primers were screened against the F1 and the recurrent parent (A. stenosperma). DNA bands present in the F1 and absent in A. stenosperma correspond to heterozygous loci and should segregate in a 1:1 ratio in the BC population. These polymorphic bands are A. cardenasii-specific (nonrecurrent parent) and were scored in the population. To be certain that polymorphic bands would correspond to heterozygous loci, and would be amplified with enough intensity to be reliably scored in the BC population (also in heterozygous state), the F1 was used during the screening step rather than the nonrecurrent parent. One hundred fifty-six primers were selected based on the size and intensity of the RAPD polymorphisms amplified. Segregating RAPD markers were identified by the manufacturer's primer code (Operon primers as A–Z, each from 1 to 20, and University of British Columbia primers as UBC 100–200, 500–600) corresponding to a particular 10-base sequence, followed by a slash and a number indicating the fragment size in base pairs. Likewise a group of framework RAPD markers was mapped in the F2 mapping population to provide additional anchor points to help align both maps.
Segregation and Linkage Analysis
A χ2 test (P = 0.05) with Yates correction factor was performed to test the null hypothesis of 1:1 segregation for all markers. Markers deviating significantly from the expected 1:1 segregation were excluded from the initial grouping and later added to the framework map as accessory markers. Linkage analysis was performed using MAPMAKER (Lander et al., 1987). All pairs of linked markers were first assigned to linkage groups using the “GROUP” command with a LOD ≥ 4.0 and recombination fraction θ = 0.30. Co-segregating markers (no recombination among markers within a linkage group) were detected from 2-point linkage data. Framework maps were constructed using only one from each set of cosegregating markers. “1st ORDER” and “COMPARE” commands in MAPMAKER were used to identify the most probable marker order within a linkage group. After establishing a framework map, the data were scanned for double cross-overs. If double cross-overs were observed adjacent to a single locus, primary data was rechecked for potential scoring errors. Additional markers were added to the framework map using the “TRY” command. Map distances in centimorgans (cM) between markers were calculated from recombination frequencies using Kosambi's mapping function (Kosambi, 1944).
Results
Primer Screening and Map Construction
Thirty-one of the 428 primers screened (7.2%) did not produce visible amplification, 145 (34%) did not detect polymorphisms between A. stenosperma and A. cardenasii, and 252 primers (58.8%) detected at least one polymorphic fragment. A total of 340 polymorphic bands, averaging nearly 1.35 polymorphic bands/primer were observed. From the 252 polymorphic primers, 156 were selected to screen the BC population. One hundred seven primers, identifying a total of 178 polymorphic loci, were reliably scored and used for mapping (1.7 mapped markers/selected primer and 0.42 mapped markers/any arbitrary primer).
At a LOD = 4.0, the map consisted of 206 markers that coalesced into 11 linkage groups. The number of mapped loci for each linkage group ranged from two to 40, comprising genetic distances of 2.5–29.3 cM (Fig. 1). Mapped markers include 167 RAPD markers, 34 Arachis root-specific cDNA clones and five Arachis shoot-specific cDNA clones. Twelve RAPD markers and two of the root-specific cDNA clones could not be assigned to any linkage group at the statistical stringency used, thus their linkage association was not determined.
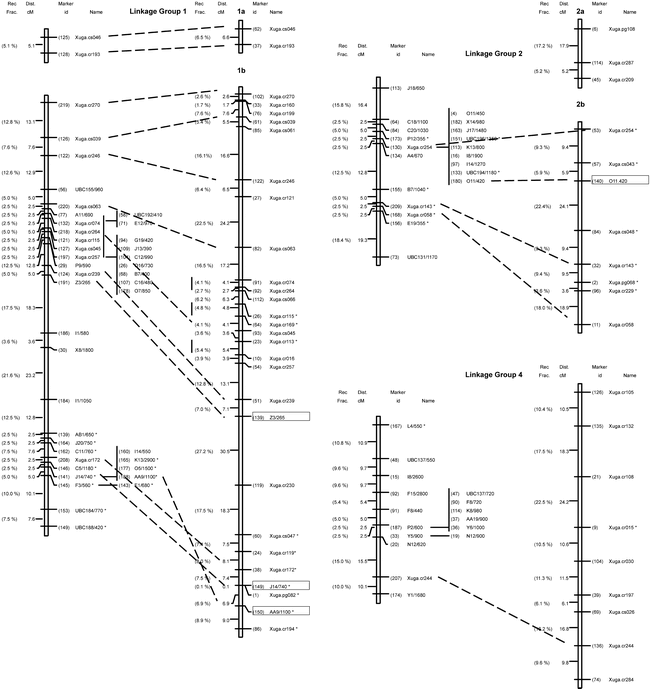
Genetic linkage maps of peanut: on left is the molecular map based in an interspecific backcross population [Arachis stenosperma × (A. stenosperma × A. cardenasii)]; on right is the RFLP map based on an F2 population derived from the same interspecific cross. Cosegregating marker loci are listed to the right of the linkage group. Loci with distorted segregation ratios from the expected 1:1 are denoted with a (.) to the right of the marker designation. Linkage groups were identified from 1 to11 according to Halward et al. (1993) based on a common set of RFLP markers, and not based on their relative size. Map distances are presented in centimorgans (Kosambi function) to the left of the linkage groups. Boxed markers represent RAPD markers segregating in a 3:1 ratio in the F2 population. Linkage groups arbitrarily numbered 9 to 11 have not been associated with the core RFLP map.
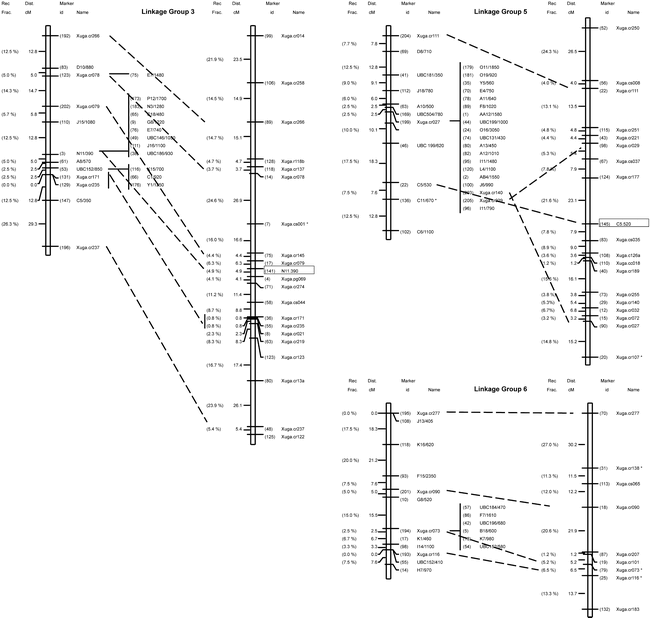
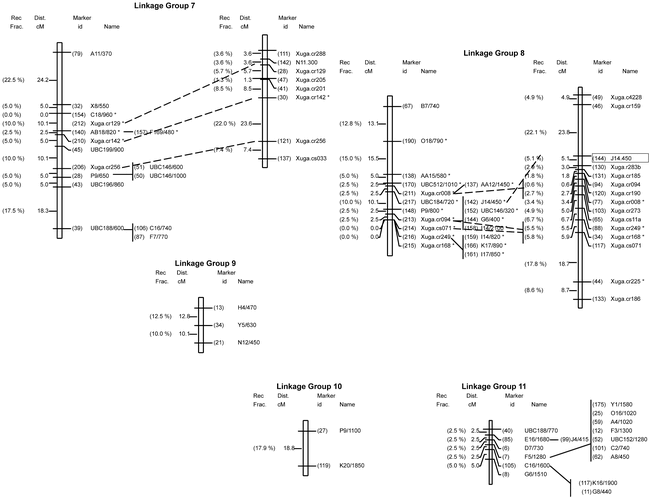
A reduction in genetic recombination compared to the F2 map was observed in all linkage groups except for group #7. The recombination fractions between identical pairs of linked markers in the F2 population studied by Halward et al. (1993) and in the BC population studied here were compared to determine whether levels of recombination observed in both mapping populations were similar. Based on two-point analysis, only five of 35 intervals (14.3%) located throughout the genome were larger in the BC map than in the F2 map (Table 1). This suggests that recombination in the F2 population exceeds that of the BC population in most regions. A more compact map, covering 800 cM (average interval 8.1 cM) was established as compared to the 1400 cM covered by the F2 map. The 39 previously mapped mapped probes and the six RAPD markers (mapped in the F2 population) were located on the same linkage groups and mostly in the same order in both populations (Fig. 1).
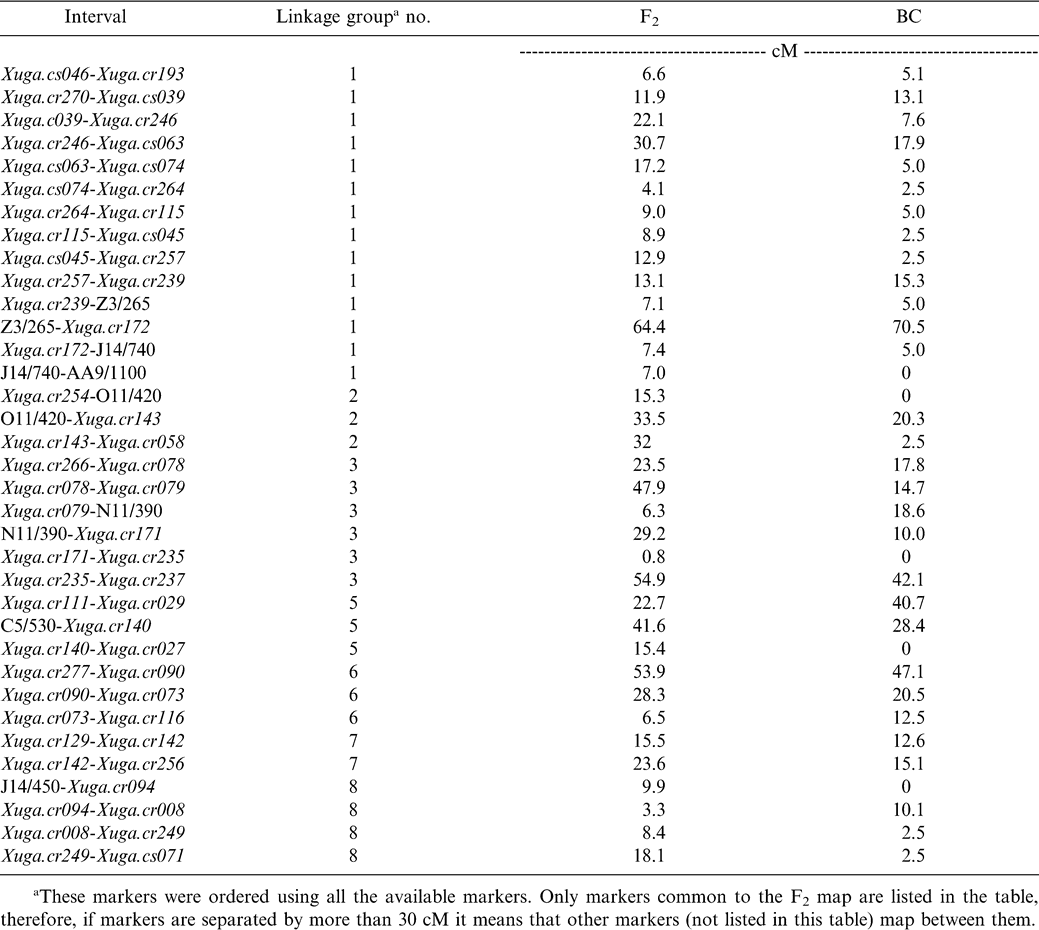
Ninety-four of the 206 mapped markers (45.6%) co-segregated into 15 clusters of different sizes in almost all linkage groups and, in most cases towards the center of the group. Clusters are formed by markers with no observed recombination. This uneven distribution of markers suggests that the frequency of recombination varies greatly from one region to another on the same chromosome. Only five RFLP markers (12.8%) were found in clusters of two or more as compared to 47% of the mapped RAPD markers (Fig. 1). The resolution was limited in several regions due to the small sample size evaluated. For example, in linkage group #5, three clones (Xuga.cr027, Xuga.cr140 and Xuga.cr029) cosegregated in the BC map but mapped to opposite ends of the linkage group in the F2 map (Fig.1). Originally, linkage group #5 in the F2 map was reported as two subgroups (5a and 5b) (Halward et al., 1993) due to the loose linkage observed between Xuga.cr177 and Xuga.cs035. The RAPD marker C5/530 mapped between the two RFLP clones and the two subgroups were joined without map expansion.
Distorted Loci
Fifty-four loci (26.2%) showed segregation distortion (a = 0.05), as is common in interspecific crosses. Eleven were RFLP (28%) and 43 (26%) were RAPD loci, suggesting that there is no prevalence of distortion with either type of marker. About one third of the distorted loci (three RFLPs and 13 RAPDs) showed an excess of the A. stenosperma alleles and all but one mapped to linkage group #2. The remaining 38 distorted loci (eight RFLPs and 30 RAPDs) showed an excess of A. cardenasii alleles and mapped to linkage groups #7, #8, and the distal end of #1.
Discussion
One of the primary uses of a genetic linkage map is to locate markers linked to genes of interest that can be either a single gene or a quantitatively inherited character. Because of the relatively low efficiency in mapping RAPDs in F2 populations due to the dominant nature of the markers, a BC population was utilized to construct a genetic linkage map of peanut. The BC map now comprises 206 markers (167 RAPDs and 39 RFLPs) distributed into 11 linkage groups.
The ratio of mapped markers per selected primer (equaling 1.7) is about half of that reported in Eucalyptus (Grattapaglia and Sederoff, 1994) and lettuce (Kesseli et al., 1994) interspecific crosses. This contrast in efficiency is probably due to differences in genomic complexity and genome size and because only polymorphic bands with an intensity score of 2 or 3 (according to Grattapaglia and Sederoff, 1994) were mapped.
A major feature of the peanut map is reduced genetic recombination, resulting in less genome coverage (800 cM) and higher clustering of markers than the F2 RFLP map of the same species (Halward et al., 1993). This higher marker density towards the middle of linkage groups could correspond to centromeric areas which are rich in repetitive DNA, thus providing a better target for RAPD markers as compared to RFLP probes (Williams et al., 1993). Reduced levels of meiotic recombination in areas surrounding the centromere also could have caused markers to cluster in these regions even though they are physically well separated. These factors, combined with the limited resolution of our mapping population with only 44 meioses, are the likely reasons for the excessive clustering and reduced recombination observed in the BC map. Hauge et al. (1993) made similar observations in Arabidopsis spp. when comparing their map with that from Reiter et al. (1992) based on a larger population size. Marker clustering was also observed by Kesseli et al. (1994) in lettuce, and by Grattapaglia and Sederoff (1994) in Eucalyptus spp.
Twenty-six percent of the loci mapped in the BC population exhibited skewed segregation ratios. This percentage was comparable to the 25% observed by Halward et al. (1993) in the peanut F2 map. This is higher than that observed by Kesseli et al. (1994), and Grattapaglia and Sederoff (1994) in lettuce, and Eucalyptus, respectively; but lower than that found by Brummer et al. (1993) in alfalfa. Distorted segregation of markers has also been reported for Lotus japonicus (Regel) K. Larsen, a model legume species (Sandal et al., 2002). The observed skewed linkage groups most likely represent real associations among loci since a chromosomal segment showing distorted segregation also will cause skewed segregation ratios in neighboring segments. The large linkage block at the distal end of linkage group #1 could really map to a different area of the genome; however, because Xuga.cr172, J14/740 and AA9/1100 map in the same position in the F2 map, it was concluded as being part of linkage group #1.
Probes that showed skewed segregation in the F2 population were also distorted in the BC population and could be mapped to the same linkage groups. Although it has been suggested that backcrosses are less sensitive to skewing than F2 populations because recombination measurements are limited to alleles coming from only one parent (Causse et al. 1994), the results with peanut showed no significant difference in the amount of distortion found in either type of mapping populations or between the RAPD and RFLP markers. The results for peanut also suggest that the cause of the distortion is not relevant to the size of the segregating population. Similar findings were reported in a common bean intraspecific F2 mapping population (Adam-Blondon et al., 1994).
The information provided by the BC map along with that provided by the F2 map can greatly aid in the identification and placement of markers linked to genes of agronomic interest into the peanut RFLP core map. Moreover, since both types of markers are known to target different areas of the genome, combining the information provided by both maps will increase the genome coverage of the current diploid peanut RFLP map.
Acknowledgements
Research was partially funded by USDA Competitive grant (#92-37300-7516) from the National Research Initiative Plant Genome Program and Peanut CRSP, USAID (grant #DAN-4048-G-SS-2065-00). Recommendations do not represent an official position or policy of the USDA or USAID.
Literature Cited
Adam-Blondon A. F. , Sevignac D. M. , and Bannerot H. 1994 A genetic map of common bean to localize specific resistance genes against anthracnose. Genome 37 : 915 – 924 .
Brummer C. E. , Bouton J. H. , and Kochert G. 1993 Development of an RFLP map in diploid alfalfa. Theor. Appl. Genet 86 : 329 – 332 .
Burow M. D. , Simpson C. E. , Starr J. L. , and Paterson A. H. 2001 Transmission genetics of chromatin from a synthetic amphidiploid to cultivated peanut (Arachis hypogaea L.): Broadening the gene pool of a monophyletic polyploid species. Genetics 159 : 823 – 837 .
Causse M. A. , Fulton T. M. , Cho Y. G. , Ahn S. N. , Chunwongse J. , Wu K. , Xiao J. , Yu X. , Ronald P. C. , Harrington S. E. , Second G. , McCough S. R. , and Tanksley S. D. 1994 Saturated molecular map of the rice genome based on an interspecific backcross population. Genetics 138 : 1251 – 1274 .
Echt C. S. , Kidwell K. K. , Knapp S. J. , Osborn T. C. , and McCoy T. J. 1993 Linkage mapping in diploid alfalfa (Medicago sativa). Genome 37 : 61 – 71 .
Fernandez M. E. , Figueiras A. M. , and Benito C. 2002 The use of ISSR and RAPD markers for detecting DNA polymorphism, genotype identification and genetic diversity among barley cultivars with known origin. Theor. Appl. Genet 104 : 845 – 851 .
Garcia G. M. , Stalker H. T. , and Kochert G. 1995 Introgression analysis of an interspecific hybrid population in peanuts (Arachis hypogaea L.) using RFLP and RAPD markers. Genome 38 : 166 – 176 .
Gill K. S. , Lubbers E. L. , Gill B. S. , Raupp W. J. , and Cox T. S. 1991 A genetic linkage map of Triticum tauschii (DD) and its relationship to the D genome of bread wheat (AABBDD). Genome 34 : 362 – 374 .
Grattapaglia D. and Sederoff R. R. 1994 Genetic linkage maps of Eucaliptus grandis and Eucaliptus urophiylla using a pseudo-testcross: Mapping strategy and RAPD markers. Genetics 137 : 1121 – 1137 .
Guyomarc'h H. , Sourdille P. , Charmet G. , Edwards K. , and Bernard M. 2002 Characterization of polymorphic microsatellite markers from Aegilops tauschii and transferability to the D-genome of bread wheat. Theor. Appl. Genet 104 : 1164 – 1172 .
Halward T. , Stalker H. T. , and Kochert G. 1993 Development of an RFLP linkage map in diploid peanut species: Applications for cultivar improvement. Theor. Appl. Genet 87 : 379 – 384 .
Hauge B. M. , Hanley S. M. , Cartinhour S. , Cherry J. M. , Goodman H. M. , Koornneef M. , Stam P. , Chang C. , Kempin S. , Medrano L. , and Meyerowitz E. M. 1993 An integrated genetic/RFLP map of the Arabidopsis thaliana genome. Plant Jour 3 : 745 – 754 .
Holbrook C. C. and Stalker H. T. 2003 Peanut Breeding and Genetic Resources. Plant Breeding Rev 22 : 297 – 356 .
Kesseli R. V. , Paran I. , and Michelmore R. W. 1994 Analysis of a detailed genetic linkage map of Lactuca sativa (lettuce) constructed from RFLP and RAPD markers. Genetics 136 : 1435 – 1446 .
Kochert G. , Halward T. , Branch W. D. , and Simpson C. E. 1991 RFLP variability in peanut (Arachis hypogaea L.) cultivars and wild species. Theor. Appl. Genet 81 : 565 – 570 .
Kosambi D. D. 1944 The estimation of map distances from recombination values. Ann. Eugenet 12 : 172 – 175 .
Krapovikas A. and Gregory W. C. 1994 Taxonomy of Genus Arachis (Legumonisae). Bonplandia 8 : 1 – 186 .
Lander E. S. , Green P. , Abrahamson J. , Barlow A. , Daly M. J. , Lincoln S. E. , and Newburg L. 1987 MAPMAKER: An interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1 : 174 – 181 .
Lin T. Y. , Kao Y. Y. , Lin R. F. , Chen C. M. , Huang C. H. , Wang C. K. , Lin Y. Z. , and Chen C. C. 2001 A genetic linkage map of Nicotiana plumbaginfolia/Nicotiana longiflora based on RFLP and RAPD markers. Theor. Appl. Genet 103 : 905 – 911 .
Paterson A. H. , Stalker H. T. , Gallo-Meagher M. , Burow M. D. , Dwivedi S. L. , Crouch J. H. , and Mace E. S. 2004 Genomics and genetic enhancement of peanut. 97 – 109 In Wilson R. F. , Stalker H. T. , and Brummer C. E. eds. Genomics for Legume Crops Amer. Oil Chemical Soc. Press .
Reiter R. S. J. , Williams J. G. K. , Feldman K. A. , Rafalski J. S. , V Tingey S. , and Skolnik P. A. 1992 Global and local genome mapping in Arabidopsis thaliana by using recombinant inbred lines and random amplified polymorphic DNAs. Proc. Natl. Acad. Sci. (USA) 89 : 1477 – 1481 .
Stalker H. T. and Mozingo L. G. 2001 Molecular markers of Arachis and marker-assisted selection. Peanut Sci 28 : 117 – 123 .
Sandal N. , Krusell L. , Radutoiu S. , Olbryt M. , Pedrosa A. , Stracke S. , Sato S. , Kato T. , Tabata S. , Parniske M. , Bachmair A. , Ketelsen T. , and Stougaard J. 2002 A genetic linkage map of the model legume Lotus japonicus and strategies for fast mapping of new loci. Genetics 161 : 1673 – 1683 .
Sun G. , Wang-Pruski G. , Mayich M. , and Jong H. 2003 RAPD and pedigree-based genetic diversity estimates in cultivated diploid potato hybrids. Theor. Appl. Genet 107 : 110 – 115 .
Ude G. , Pillay M. , Ogundiwin E. , and Tenkouano A. 2003 Genetic diversity in an African plantain core collection using RAPD and AFLP markers. Theor. Appl. Genet 107 : 248 – 255 .
Williams J. G. K. , Hanafey M. K. , Rafalski J. A. , and Tingey S. V. 1993 Genetic analysis using random amplified polymorphic DNA markers. Methods in Enzymol 218 : 704 – 740 .
Williams J. G. K. , Kubelik A. R. , Livak K. L. , Rafalski A. J. , and Tingey S. V. 1990 DNA polymorphism amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res 18 : 6531 – 6535 .
Yu G. X. and Wise R. P. 2000 An anchored AFLP- and retrotransposon-based map of diploid Avena. Genome 43 : 736 – 749 .