

Introduction
Pod rot is the most consistent and frequent disease problem in the west Texas peanut producing area. Pod rot can be caused by a number of different fungi including Rhizoctonia solani, Pythium spp., Sclerotium rolfsii, and Fusarium spp. (Porter et al., 1982). A survey of chemical usage on peanuts in the southwestern U.S. (Smith et al., 1998) indicated that most of the fungicides used in Texas were active against R. solani, but not against Pythium spp. However, beginning in 1999, Pythium was isolated more frequently from pod rot samples sent to the Texas A&M University Agriculture and Extension Center - Lubbock than was Rhizoctonia (T. Wheeler, unpublished). There is, therefore, a need to clarify the relative frequency of both Rhizoctonia and Pythium in association with pod rot in this region.
Fungicides with activity against Pythium spp. and recommended for pod rot control or suppression include mefenoxam and azoxystrobin (Texas Peanut Production Guide, 2001). Mefenoxam is a stable compound that is resistant to a wide range of pH, temperature, and light conditions (Sukul and Spiteller, 2000). When mefenoxam performs poorly it may be due to either biodegradation (Sukul and Spiteller, 2000) or fungal resistance (Davison and McKay, 1999). Insensitivity to metalaxyl by a Pythium species was first reported on turfgrass to P. aphanidermatum (Sanders, 1984). However, insensitivity to mefenoxam from other Pythium species has been reported only rarely (Moorman and Kim, 2004; Moorman et al., 2002; Taylor et al., 2002). In west Texas, mefenoxam has been used widely as a cotton seed treatment, but there has been relatively little use of mefenoxam in peanut production (Smith et al., 1998). Biodegradation of mefenoxam is associated with extensive usage (Davison and McKay, 1999) and is probably not a factor.
Azoxystrobin has activity against a wide range of fungi, but oomycetes are generally less sensitive than the Ascomycetes, Dueteromycetes, or Basidiomycetes (Clough and Godfrey, 1998). Azoxystrobin is an effective and economical tool for managing mixtures of pathogens causing pod rots, particularly when both Rhizoctonia and Pythium spp. are involved. Fungal insensitivity to azoxystrobin has been documented in many fungi (Vinelli and Dixon, 2002; Wong and Wilcox, 2002; Mavroeidi and Shaw, 2005). An understanding of the sensitivity of Pythium spp. isolated from peanuts to azoxystrobin and mefenoxam is important for pod rot management in fields where pathogenic Pythium spp. are present.
Materials and Methods
Survey
Peanut fields (107) were selected without bias as to diseases in Cochran, Crosby, Dawson, Gaines, Hockley, Lubbock, Terry, and Yoakum counties in the Texas Southern High Plains. Fields were identified by driving through multiple (paved and dirt) roads in a county and marking the presence of peanut fields, and by flying over counties and marking the presence of peanut fields. Each field in a county received an identification number; then a series of random numbers were drawn and the subset of fields to be surveyed was selected. Each field was sampled in an identical manner regardless of size. Fields ranged from 12 to 99 ha. Sampling began in the last week of August and continued throughout September of 2003. Fields sampled included Runner, Spanish, Virginia, and Valencia types. Varieties in some fields could be identified and included Flavorunner 458 (most frequent), Florunner, NC7, Spanco, Tamspan 90, Valencia 101, VC2, and Viraguard. Maturity at the time of sampling ranged from only small, immature pods all the way to mature, ready for harvest pods.
Four transects were made across each field, with 10 locations sampled per transect. The sample location was selected by pacing even distances along the transect. At each sample location, approximately 1m of plants in a row were pulled up and all the discolored pods were collected. The soil underneath the plants was searched for any pods that had been dislodged from the plants. Pods with any discoloration in the 40 sample locations of a field were placed in a paper bag and refrigerated until the sample was processed. Processing (conducted within 1 wk of sampling) consisted of examining each pod for necrosis, weighing all necrotic pods, and selecting up to 16 pods for assay. In some cases, there were less than 16 pods from a field with necrosis. Pods selected for assays represented the range of symptoms expressed for a field (black, brown, white, rotted, and sunken type lesions) were washed, dried, and two small pieces (approximately 0.5 to 1 cm) were plated out on water agar and a Pythium selective medium (Lewis and Filonow, 1990). Plates were incubated at room temperature (21 to 23C) and monitored daily for mycelial growth, for seven days. Mycelia growing from these pod pieces were transferred to potato dextrose agar and identified to the genus level. Twenty-six of 85 isolates identified as belonging to the genus Pythium (no more than 4 isolates from a given field) were sub-cultured on corn meal agar (CMA) + 10 µg rifampicin/ml, and identified to species based on the descriptions from Plaats-Niterink (1981). There were 50 unidentified Pythium isolates that did not produce any structures that were useful for taxonomic purposes and nine identified as the nonpathogenic P. oligandrum. Pythium isolates that were selected for species identification represented all the fields where isolates were found, that did not quickly become contaminated by bacteria. The pathogens present within each field were divided into four classes: 1. neither Rhizoctonia nor Pythium present; 2. Rhizoctonia present without Pythium; 3. Pythium present without Rhizoctonia; or 4. Rhizoctonia and Pythium both present.
Fungicide Sensitivity
Isolates of Pythium spp. were maintained on CMA + 10 µg rifampicin/ml. One cm2 plugs of mycelium were placed on CMA plates containing the following concentrations of mefenoxam : 0, 0.0096, 0.096, 0.96, and 9.6 µl/ml of medium. The source of mefenoxam was Ridomil Gold EC (47.6% ai, Syngenta Crop Protection, Greensboro, NC). The size of the colony was measured approximately 40 hrs later. Percent inhibition was expressed as a percentage of the colony diameter in the absence of mefenoxam (Ridomil Gold EC). Plates were grown on a laboratory bench at room temperature which varied from 18–23 C. The plates were not kept in the dark, since mefenoxam is resistant to sunlight (Sukul and Spiteller, 2000). Each isolate was tested on a given date with five concentrations of fungicide (including the 0 concentration) and three replications per isolate/concentration, arranged in a randomized complete block design. Each isolate was tested at least two times. Isolates were run in subgroups from November 03 through February 04.
Azoxystrobin was tested in a fashion similar to that of mefenoxam, except that higher concentrations were used (10 µl, 100 µl, 1,000 µl, and 4,000 µl/ml). The source of azoxystrobin was Abound FL (22.9% ai, Syngenta Crop Protection, Greensboro, NC).
The concentration that effectively reduced colony diameter by 50% (EC50) was estimated for all isolates. A number of different models were tested to estimate EC50. However, the natural log transformation of both fungicide concentration and percentage inhibition of fungal growth provided the most consistent fit for data.
Results and Discussion
Survey
Both Pythium and Rhizoctonia spp. were isolated from rotted pods in similar percentages. The classes representing Rhizoctonia alone, Pythium alone, and Rhizoctonia and Pythium together were found in 19, 23, and 16% of the fields surveyed, respectively. In 42% of the fields, neither Rhizoctonia nor Pythium were found. In those cases, either no pod rot was identified, or only superficial necrosis was present which is typical of the black hull fungus Thielaviopsis basicola. Thielaviopsis basicola was not isolated from the rotted pods, but would have required different selective media. Sclerotium rolfsii was isolated from pods in 4% of the fields. Fusarium was isolated from pods in all fields. The overall condition of the pods was good, but when there was substantial rot, then typically either Rhizoctonia or Pythium were isolated. The 2003 summer was dry, particularly during the time of pod formation, so the environment was not conducive for pod rot. All of the surveyed peanut fields were irrigated to some degree. The frequency of Pythium and Rhizoctonia spp. isolated from rotted pods in a survey can change from year to year (Hollowell et al., 1998). In North Carolina, both Pythium spp. and Rhizoctonia spp. were the two pathogens most frequently isolated in a survey of peanut fields, but Rhizoctonia spp. predominated in one year, and Pythium spp. predominated in two years of a three year survey (Hollowell et al., 1998). It is possible that the pod rot percentage in west Texas will increase in years where the environment is more conducive for disease, and that the relative percentages of disease due to Pythium and Rhizoctonia may change. However, it is clear that this region has pod rot associated with both Rhizoctonia spp. and Pythium spp., although pod rot overall during a dry summer is relatively low.
Pythium oligandrum was frequently isolated from rotted peanut pods, but is not considered a plant pathogen (Benhamou, et al., 1999). Twenty-six Pythium isolates were identified to the species level, not including P. oligandrum. The species of Pythium most frequently isolated in west Texas were P. ultimum (9 isolates), P. myriotylum (10 isolates), and P. irregulare (6 isolates). One isolate was identified tentatively as P. heterothallicum. In a North Carolina survey, P. irregulare and P. spinosum were the species most frequently identified (Hollowell et al., 1998). Other species identified in North Carolina on peanuts were P. myriotylum, P. dissotocum, P. ultimum, P. vexans, P. dimorphum, and P. paroecandrum. The “classic” Pythium species on peanut is considered to be P. myriotylum (Porter et al., 1982).
Fungicide Sensitivity
The pathogenic Pythium isolates (those associated with pod rot, but not including P. oligandrum) had EC50 which ranged from 0.001–0.270 µl ai mefenoxam/ml. Only one isolate was identified as P. heterothallicum, and it was quite sensitive to mefenoxam (EC50 = 0.011 µl ai of mefenoxam/ml). Five isolates of P. irregulare that were obtained from three fields had EC50 that ranged from 0.049–0.173 µl ai of mefenoxam/ml, with an average EC50 of 0.119 µl/ml. Seven isolates of P. myriotylum that were obtained from five fields, had EC50 that ranged from 0.012–0.163 µl ai of mefenoxam/ml and averaged 0.074 µl/ml. Three isolates of P. oligandrum from three fields had EC50 that ranged from 0.008–0.099 µl ai of mefenoxam/ml, and averaged 0.042 µl/ml. Two isolates of P. ultimum from two fields had EC50 of 0.005 and 0.006 µl ai of mefenoxam/ml. Within the same field, isolates of Pythium could have very similar EC50 (G31A, B, and C ranged from 0.001 to 0.008 µl ai of mefenoxam/ml), or very different EC50 (L3A and L3C were 0.033 and 0.270 µl ai of mefenoxam/ml, respectively, Table 1). None of the isolates would be considered resistant to mefenoxam. Mefenoxam resistance has been defined as EC50>100 µl/ml (Moorman et al., 2002; Taylor et al., 2002) and the least sensitive isolate (L3C) had an EC50 of 0.27 µl/ml. The EC50 of P. ultimum var. sporangiiferum from sugar beet ranged from 0.050–1.3 µl/ml (Branter and Windels, 1998). The EC50 of P. irregulare obtained from apple roots was much higher (0.200–0.790 µl/ml) than from P. ultimum (0.010–0.170 µl/ml) (Mazzola et al., 2002). Pythium irregulare that was obtained from plants taken from commercial greenhouses (a mixture of plant species) was frequently (21 of 57 isolates) resistant to mefenoxam, while no resistant P. myriotylum, and only 1 of 13 resistant P. ultimum isolates were found (Moorman et al., 2002). The sensitivity of mefenoxam in the Pythium isolates examined could be attributed to their lack of exposure to this fungicide. Previous exposure was probably limited to cotton seed treatments, which would have placed minimal selection pressure on Pythium populations.
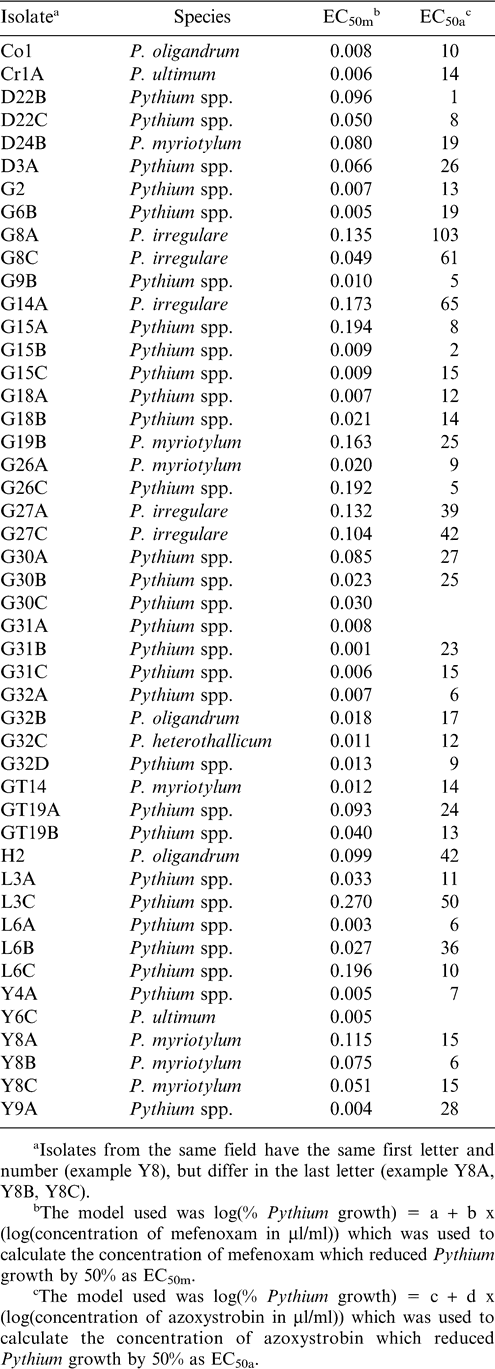
Sensitivity of Pythium spp. to azoxystrobin was much lower than to mefenoxam. However, some fungi use an alternative respiratory pathway that can interfere with the activity of azoxystrobin in a petri dish assay (Stevenson et al., 2004). This means that these results cannot be compared directly to assays where salicylhydroxamic acid is used to inhibit Pythium from using an alternative respiratory pathway, though no published in vitro studies with azoxystrobin and Pythium were found. Sensitivity of different Pythium isolates to azoxystrobin in this study ranged from 1 to 103 µl ai of azoxystrobin/ml. Pythium heterothallicum had an EC50 of 12 µl ai of azoxystrobin/ml. The EC50 for P. irregulare ranged from 39–103 µl ai of azoxystrobin/ml and averaged 62. The EC50 for P. myriotylum ranged from 5–25 µl ai of azoxystrobin/ml and averaged 14.7. The EC50 for P. oligandrum ranged from 10–42 µl ai of azoxystrobin/ml and averaged 23. The EC50 for P. ultimum was 14 for one isolate and no acceptable model could be fitted to the second isolate (Y6C). Isolate Y6C had a slight decrease in growth from 10 µl to 1,000 µl azoxystrobin/ml and no growth occurred at 4,000 µl azoxystrobin/ml. All the isolates of P. irregulare had less sensitivity to azoxystrobin than any of the other pathogenic Pythium isolates that were identified to species. Of the isolates that were not identified to species, the most sensitive P. irregulare isolate (G27A) was less sensitive than 93% of the remaining isolates.
Summary and Conclusions
Pythium spp. are an important component of the pod rot complex in west Texas. Chemical management options for this organism include mefenoxam (control) and azoxystrobin (suppression). There do not appear to be resistance issues with respect to mefenoxam use on peanut. There is evidence in other crops and with certain fungal species, that resistance to azoxystrobin is associated with increased usage, and especially with low dosages of the product (Gisi et al., 2000). The majority of azoxystrobin treatments are applied to control Rhizoctonia pod rot, and the recommended rate for this fungus is 37.4–74.9 g ai/ha of azoxystrobin. Typically, producers will use 50–56 g ai/ha for Rhizoctonia control (broadcast rate). This may be a sublethal rate to control Pythium, for which the required rate is 74.9 g ai/ha. Continued reliance on azoxystrobin at sublethal rates for Pythium spp. will probably lead to more isolates with reduced sensitivity to this fungicide.
Acknowledgements
Funding was provided by the Texas Agricultural Experiment Station, USDA-ARS, and the Texas Peanut Producers Board, and products were donated by Syngenta Crop Protection.
Literature Cited
Benhamou N. , Rey P. , Picard K. , and Tirilly Y. 1999 Ultrastructural and cytochemical aspects of the interaction between the mycoparasite Pythium oligandrum and soilborne plant pathogens. Phytopath 89 : 506 – 517 .
Brantner J. R. and Windels C. E. 1998 Variability in sensitivity to metalaxyl in vitro, pathogenicity, and control of Pythium spp. on sugar beet. Plant Dis 82 : 896 – 899 .
Clough J. M. and Godfrey C. R. A. 1998 The strobilurin fungicides. 109 – 148 In Hutson D. and Migamoto J. eds. Fungicidal Activity: Chemical and Biological Approaches to Plant Protection Wiley and Sons West Sussex, England .
Davison E. M. and McKay A. G. 1999 Reduced persistence of metalaxyl in soil associated with its failure to control cavity spot of carrots. Plant Pathology 48 : 830 – 835 .
Lewis P. I. and Filonow A. B. 1990 Reaction of peanut cultivars to Pythium pod rot and their influence on populations of Pythium spp. in soil. Peanut Sci 17 : 90 – 95 .
Gisi U. , Chin K. M. , Knapova G. , Farber R. K. , Mohr U. , Parisi S. , Sierotzki H. , and Steinfeld U. 2000 Recent developments in elucidating modes of resistance to phenylamide, DMI and strobilurin fungicides. Crop Prot 19 : 863 – 872 .
Hollowell J. E. , Shew B. B. , Beute M. K. , and Abad Z. G. 1998 Occurrence of pod rot pathogens in peanuts grown in North Carolina. Plant Dis 82 : 1345 – 1349 .
Mavroeidi V. I. and Shaw M. W. 2005 Sensitivity distributions and cross-resistance patterns of Mycosphaerella graminicola to fluquinconazole, prochloraz and azoxystrobin over a period of 9 years. Crop Prot 24 : 259 – 266 .
Mazzola M. , Andrews P. K. , Reganold J. P. , and Andre Levesque C. 2002 Frequency, virulence and metalaxyl sensitivity of Pythium spp. isolated from apple roots under conventional and organic production systems. Plant Dis 86 : 669 – 675 .
Moorman G. W. and Kim S. H. 2004 Species of Pythium from greenhouses in Pennsylvania exhibit resistance to propamocarb and mefenoxam. Plant Dis 88 : 630 – 632 .
Moorman G. W. , Kang S. , Geiser D. M. , and Kim S. H. 2002 Identification and characterization of Pythium species associated with greenhouse floral crops in Pennsylvania. Plant Dis 86 : 1227 – 1231 .
Plaats-Niterink A. and Van Der J. 1981 Monograph of the genus Pythium. In Gams G. W. and Jacobs R. P. W. M. eds. Studies in Mycology No. 21 Centraalbureau voor Schimmelcultures Baarn, Netherlands .
Porter D. M. , Smith D. H. , and Rodriguez-Kabana R. 1982 Peanut plant diseases. 326 – 410 In Pattee H. E. and Young C. T. eds. Peanut Science and Technology Proc. Amer. Peanut Res. Educ. Soc Yoakum, TX .
Sanders P. L. 1984 Failure of metalaxyl to control Pythium blight on turfgrass in Pennsylvania. Plant Dis 68 : 776 – 777 .
Smith D. T. , New N. G. , and Criswell J. T. 1998 Pests, pesticide use and management practices in the peanut industry in the Southwestern U. S., Dept. Technical Report 98-08, Texas AM University System.
Stevenson K. L. , Langston D. B. , and Seebold K. W. 2004 Resistance to azoxystrobin in the gummy stem blight pathogen documented in Georgia. Online. Plant Health Progress doi:10.1094/PHP-2004-1207-01-RS.
Sukul P. and Spiteller M. 2000 Metalaxyl: persistence, degradation, metabolism, and analytical methods. Rev. Environ. Contam. Toxicol 164 : 1 – 26 .
Taylor R. J. , Salas B. , Secor G. A. , Rivera V. , and Gudmestad N. C. 2002 Sensitivity of North American isolates of Phytophthora erythroseptica and Pythium ultimum to mefenoxam (metalaxyl). Plant Dis 86 : 797 – 802 .
Texas Peanut Production Guide 2001 Texas Cooperative Extension. B-1514. http://lubbock.tamu.edu/peanut/docs/PeanutProdGuide2001.pdf .
Vincelli P. and Dixon E. 2002 Persistence to Q(0) fungicides in isolates of Pyricularia grisea from perennial ryegrass. Plant Dis 86 : 235 – 240 .
Wong F. P. and Wilcox W. F. 2002 Sensitivity to azoxystrobin among isolates of Uncinula necator: baseline distribution and relationship to myclobutanil sensitivity. Plant Dis 86 : 394 – 404 .