

INTRODUCTION
Global peanut production was valued at approximately $47 billion in 2020, with over $1 billion attributed to the U.S. crop. (FAOSTAT, 2020). It is the third major oilseed of the world behind soybean [Glycine max (L.) Merr.] and cotton (Gossypium hirsutum L.). Peanuts are high in protein, fiber, tocopherols, unsaturated fatty acids, and other nutritionally advantageous compounds (Ros, 2010), and are used for oil extraction, edible food products and confectionary purposes across the globe. Sustainable and profitable peanut production in the U.S. is maintained by the release of improved disease-resistant cultivars, but resistance to all threatening pathogens is not currently available.
Peanut smut, a soilborne fungal disease caused by Thecaphora frezzii, was first described by Carranza and Lindquist (1962) infecting wild peanut in Brazil. The disease did not appear in commercial peanut fields until 1995, when it was reported in Argentina (Marinelli et al., 1995). Pods of plants infected by T. frezzii may be hypertrophied (Cazón et al., 2018). Colonized cells in the seed pods are deformed and replaced with a smutted mass (sorus) formed by teliospores, which are the inoculum source for the disease (Rago et al., 2017). Once in the soil, spores of the pathogen have been observed to remain infective for more than four years (Cazón et al., 2016). Thecaphora frezzii spores are easily spread by wind and weather events and the spread of the pathogen is sporadic and unpredictable. Symptoms of peanut smut infection are not visually apparent on plant foliage above ground but are revealed upon digging and shelling of peanut pods. Furthermore, pathogen spores may be concealed in peanut seed (Cazón and Rago, 2016), making visual detection less accurate.
Peanut smut has spread throughout Argentina and is now reported to be in 100% of Argentine peanut-producing fields. Disease incidence and severity varies with location but yield reductions as high as 51% have been reported (Cazón et al., 2018; Rago et al., 2017). Integrated pest management of the peanut smut has been limited. Ganuza et al. (2017) reported that treatment with the biocontrol agent Trichoderma harzianum ITEM 3636 may reduce peanut smut severity by up to 25%. Fungicidal management of the pathogen has proven largely ineffective. However, a report discussing a mitogenome of T. frezzii identified possible fungicide target genes (Arias et al., 2019), and a recent study indicated that cyproconazole or azoxystrobin application can reduce incidence significantly (Parades et al., 2021), particularly in nanocrystal-based formulations (Camiletti et al., 2021).
Host-plant resistance to peanut smut should provide sustainable protection against the disease. Resistance has been discovered among several wild Arachis species and landraces (Bressano et al., 2019; de Blas et al., 2019), and attempts to introgress resistance to smut from wild to cultivated peanut have been made (Bressano et al., 2019). More recently some accessions in the USDA peanut mini-core collection (Holbrook and Dong, 2005) were screened for phenotypic reaction to smut infection (Wann et al., 2020). Peanut cultivars released in the past have shown only moderate resistance to smut (Farías et al., 2011; Oddino et al., 2013). A recent release in Argentina demonstrates resistance ( https://www.criaderoelcarmen.com.ar/es/variedades.html), and the agronomic performance of that cultivar is still being investigated.
The rapid spread of this disease and lack of control measures have caused great concern in Argentina and within peanut research and production communities worldwide. Although peanut smut has not been reported in cultivated peanut outside of Argentina, proactive measures must be taken to protect the global peanut industry. Therefore, the objective of this research was to identify sources of smut resistance by screening purified peanut germplasm from the USDA peanut mini-core collection along with selected U.S. peanut cultivars and breeding lines.
Materials and Methods
Site Description. A smut screening test was carried out across 3 seasons (2017-2020) in fields adjacent to a farm (32°45'03.6"S 63°45'56.3"W) in the town of General Deheza (Córdoba Province, Argentina) under rainfed conditions. Soil at the site was a General Deheza coarse-silty sandy loam type (0.3% slope, coarse-silty, mixed, thermic, Entic Haplustoll). The test was planted one kilometer downwind from a peanut processing plant where peanut smut is prevalent. The production field site was on a rotation schedule of Zea mays L.-Zea mays L.-Glycine max (L.) Merr-Arachis hypogaea L. (corn-corn-soybean-peanut), where the preceding crop of the test site was always soybean.
Plant Material. Plant materials screened in this study included 208 peanut genotypes including accessions from the USDA peanut germplasm collection, U.S. peanut breeding lines, and released U.S. cultivars (Table 1). All entries/genotypes were purified by single-seed descent (SSD) in greenhouses and seed was increased from each purified genotype prior to shipment and testing. Seeds of each entry were packaged and shipped from the U.S. to Argentina approximately two months prior to planting. Remnant seed was kept in the U.S. and stored at 4 ͦ C as potential sources of smut resistance in breeding programs. Entries demonstrating ≤ 10% disease incidence were retained for testing the following year, but all others were discarded. Therefore, the number of years each entry was tested varied from one to three (Table 1).

Complete listing of peanut entries tested for resistance to peanut smut during 2017-2020, along with number of years tested, total pods tested per year, and percent disease incidence per year.
Field Experimental Design. The cumulative test (2017-2020) included 208 experimental entries and two control entries (one resistant and one susceptible). Control entries were planted each year while the evaluated entries varied in number, as mentioned previously (Table 1). The control entries were replicated a different number of times depending on the season. In 2017, the replications (four blocks) were split, and the first block contained the resistant control five times and the susceptible four times, along the diagonal of the block. In the second block, the resistant control was present five times and the susceptible five times. Finally, the third block contained the resistant control five times and the susceptible four times. In 2018 , the control entries were planted along three diagonal transects of the plot, with 23 replications of the resistant and 22 replications of the susceptible entries. In 2019 the design was similar to the previous season, with 3 diagonals containing alternating resistant and susceptible controls. The resistant control was present 26 times and susceptible was present 25 times. As the previous season, both were also included as entries with 3 reps for each.
Prior to planting, soil samples were taken, and quantitation of disease pressure was performed as described by Marinelli et al. (2008). The tests were planted on 18 Dec. 2017, 7 Nov. 2018, and 23 Dec. 2019, using an augmented grid design with three replications, using single plants as experimental units (one plant per plot). Plots were spaced one m apart on all sides. Plots were managed for weeds and diseases following the extension guidelines of the Instituto Nacional de Tecnología Agropecuaria (INTA).
Harvest and Evaluation of Smut Resistance. Plants were dug 120, 147, and 140 days after planting in 2018, 2019, and 2020, respectively. Each pod was opened by hand and the presence or absence of T. frezzii sori or spore masses on the kernels of each pod was recorded. The smut incidence for each plot (as described below) was considered as the evaluation for smut reaction. Each replication was evaluated by a different person and the same evaluators were employed over the three seasons.
Statistical Analysis. To determine the inter- evaluator reliability among smut evaluations performed by different persons, all the pods from 3 plots (replications) belonging to 10 genotypes of unknown smut reaction were hand-opened and subsequently displayed over a board in an ordered manner so that each pod corresponded to an ordered number. Five persons took turns individually evaluating each pod for the presence or absence of smut. In this way, it was possible to assign 5 evaluations for each numbered pod. Based on these 5 ratings, it was possible to do an inter-evaluator reliability assessment. Interrater reliability was calculated from 5 evaluators and 2,803 individual pods using Fleiss' Kappa in R (RStudio Team, 2020). Percentage of Smut Incidence (%SI) in a plot was calculated as %SI=[(number of infected pods/total number of pods)*100].
Analysis of variance (ANOVA) was used to ascertain differences among entries for percent smut incidence using PROC MIXED in SAS Version 9.4 (SAS Institute, 2013). To determine difference among all entries tested, each year was analyzed separately because the entries evaluated differed each year due to removal of susceptible genotypes. Since standard errors differed, generation of a single LSD value was not possible, and treatment mean letter grouping was accomplished by using the SAS macro PDMIX800 (Saxton, 2000). Analysis was performed across years for those entries tested (28 plus two controls) for three consecutive years (Table 3), where year was included as a random effect. Protected post-hoc pairwise comparisons were conducted on the least square means (DIFF option in an LSMEANS statement). A significance level of P < 0.05 was used for the pairwise comparisons. To seek association between collection origin and/or growth habit with disease incidence, analysis of variance was performed to assess disease differences for the four continents of collection origin.
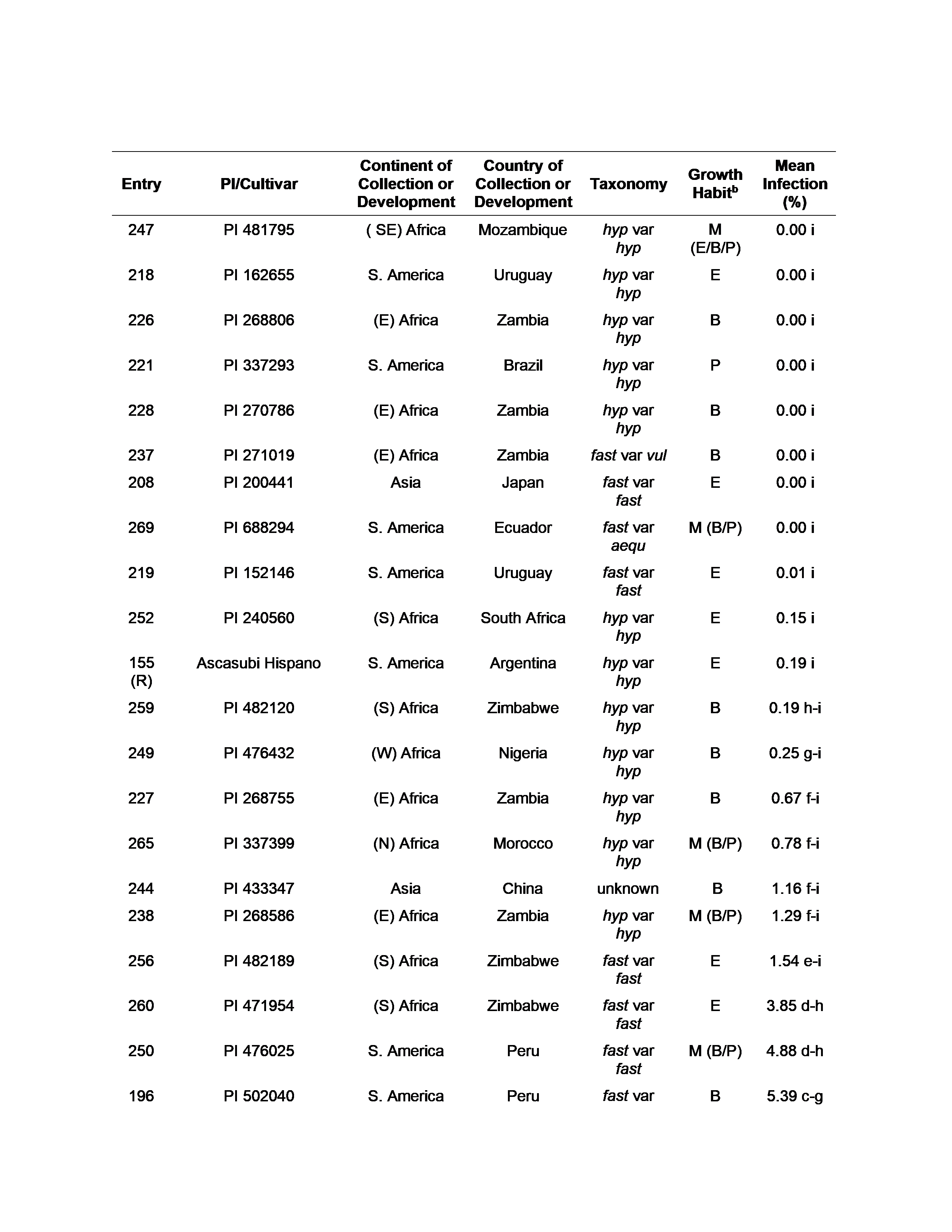
U.S. peanut germplasm evaluated for three years, 2017-2020, near General Deheza, Córdoba Province, Argentina, exhibiting ≤ 10% incidence of peanut smut, including susceptible (entry 160) and resistant (entry 155) controls.a
Results and Discussion
The peanut growing season in Argentina roughly spans the months of November through May. Field plots were not irrigated, and the yearly rainfall total varied over the 2017-2020 period of testing. In 2017-2018, the total yearly rainfall at the testing site was 255 mm, as the region experienced a severe drought. Rainfall during the 2018-2019 and the 2019-2020 seasons was much closer to the average (605 mm) for the region at 706 mm and 614 mm, respectively. Soilborne inoculum at the testing site was consistent over the testing period at 5790 (± 1006), 6100 (± 1169), and 5650 (± 1307) teliospores/g soil, in 2017-2018, 2018-2019, and 2019-2020, respectively (Table 2).

Thecaphora frezzii spore count (disease pressure) in testing sites near General Deheza, Argentina, 2017-2020.
There was strong agreement among scores given by the five evaluators, evidenced by a Kappa value of 0.868 (P < 0.001). A value of 0.454 was obtained when Kappa was calculated using the 554 pods that were evaluated as infected by at least 1 evaluator, indicating that the 5 evaluators were in reasonable agreement as to when a pod was infected. The average number of pods per entry varied by year (Table 1). A total of 106 entries were tested in 2017-2018, along with accepted resistant and susceptible controls. The observed incidence of smut ranged from 0-75%, with a mean incidence of 16.4%. The resistant control, Ascasubi Hispano ( https://inta.gob.ar/variedades/ascasubi-hispano) averaged 0.3% incidence while the susceptible control, PI 386349 (Colorado Irradiado INTA) averaged 24.1%. Significant differences in disease incidence were noted among entries. Thirty-five (33%) of the entries averaged less than 10% incidence and were retained for further testing the following year. The range of smut incidence for most entries fell between those of the susceptible and resistant controls. Thirteen entries were statistically similar in resistance to the accepted resistant control Ascasubi Hispano, and 7 entries exhibited extreme susceptibility with incidences statistically different and above that of the accepted susceptible control Colorado Irradiado INTA.
In the 2018-2019 growing season, 103 entries (including 35 retained from the prior year) were tested and smut incidence ranged from 0-37.9%, with a mean incidence of 13.5%. The resistant and susceptible controls averaged 0.4 and 34.9% smut incidence, respectively. Significant differences in disease incidence were again noted among entries. Forty-four (43%) of the entries averaged less than 10% incidence and were retained for further testing the following year. Resistance was confirmed for a second year for 27 of the 35 entries retained from testing in 2017-2018, and these were retained for a third year of testing. The range of smut incidence for most entries fell between the range of the susceptible and resistant controls.
Eighty-five entries were tested in 2019-2020, including 44 retained from the prior year. Observed smut incidence ranged from 0-39.7%, with a mean incidence of 6.9%. Resistant and susceptible controls averaged 0 and 16.3% disease, respectively. Significant differences in reaction to smut were again noted among entries. A third year of resistance (less than 10% incidence) was confirmed for 25 of the 44 entries retained from the prior year and a second year of resistance was confirmed for 12 entries. An additional 16 entries exhibiting less than 10% smut incidence were identified and those having less than 5% disease were retained for testing outside the scope of this study.
According to the Germplasm Resources Information Network (GRIN, 2021), entries used in this study were collected from or developed in four continents, (1) Asia, (2) Africa, (3) South America, and (4) North America. Analysis of variance performed to determine if an association between disease incidence and collection origin exists generated an overall p-value of 0.09 and did not warrant a pairwise comparison. Although there did appear to be some natural separation between these continents, no significant association between collection origin and disease susceptibility was found.
Twenty-five entries tested over a three-year period averaged less than 10% disease incidence (Table 3). Among those 25 entries, eight entries appeared immune to smut, averaging 0% infection. Taxonomy information for the immune group of entries (taken from GRIN) revealed that 5 of the 8 entries belonged to A. hypogaea subsp. hypogaea var hypogaea. The other three immune entries were of A. hypogaea subsp. fastigiata, with one each of var aequatoriana , var fastigiata, and var vulgaris. Examination of growth habit information, also taken from GRIN, showed that all growth habits (erect, bunch, spreading or prostrate) were represented among entries with 10% or less disease incidence, including the immune entries. According to GRIN, many entries were reported to have ‘mixed’ growth habit, indicative of seed that may have been pooled upon collection. In short, no significant correlation between disease incidence and origin, growth habit, or taxonomy was found for the entries tested in this study.
To date, all commercial cultivars planted widely in Argentina are susceptible to T. frezzii infection, which has contributed to the rapid spread of this pathogen through the growing region of Cordoba (Cignetti et al., 2010). Without effective chemical means of control, identification, and incorporation of resistance genes to develop cultivars with host-plant resistance remains the best option for peanut smut management. Sources of resistance in wild Arachis species and landraces have been identified (de Blas et al., 2019; Oddino et al., 2017; Soave et al.,2014) and some have been introgressed into a cultivated, tetraploid background (Bressano et al., 2019; de Blas et. al., 2019) in a stable and heritable manner. However, reports of smut resistance occurring naturally in cultivated, tetraploid backgrounds are scarce.
Wann et al. (2020) recently evaluated peanut germplasm, including 97 peanut accessions from a purified U.S. peanut mini-core collection (Chen et al., 2014), for reaction to T. frezzii in infested fields near General Cabrera, also in Córdoba Province, Argentina. Disease incidence ranging from 0-23% was observed, and twelve accessions tested from the U.S. peanut mini-core collection had no incidence of T. frezzii infection. The results obtained in our study support those findings in part. The genotypes evaluated in this study include 81 of the 97 mini-core plant introductions (PIs) examined by Wann et al. Of the accessions present in both studies, only two (PI 268806 and PI 481795) exhibiting zero disease by Wann and colleagues appeared immune in our study. However, four accessions reported with no smut incidence by Wann et al. (PIs 268696, 337399, 482120, and 494018) were highly resistant (< 1% disease), but not immune in the trials conducted in this study.
The germplasm evaluated in this study were not from the collection of Chen et al. (2014) which was purified using phenotypic characteristics. Instead, each mini-core accession used in our study was purified from seed provided by the USDA National Plant Germplasm Repository System in Griffin, Georgia using SSD. This difference in germplasm source may explain some differences between these results with those reported by Wann and colleagues (2020). A genetic comparison of our purified accessions with those developed by Chen et al. (2014) would test this hypothesis. Other possible explanations for the differences between the studies include differing test locations, disease pressures, environmental conditions, and experimental design.
Of the 34 breeding lines and cultivars examined in this study, TifNV-High O/L (Holbrook et al., 2017) demonstrated moderate resistance to T. frezzii infection, at 6.7% over 3 years. Within the pedigree of TifNV-High O/L, the background of ancestor C99-R (Gorbet and Shokes, 2002) contains two PIs (203396 and 259785) that may have contributed to smut resistance. Neither of these PIs were evaluated for smut resistance here nor by Wann et al. (2020). The ancestry of TifNV-High O/L also includes TxAG-6, an amphidiploid resulting from a [(A. cardenasii x A. diogoi) x A. batizocoi] cross and subsequent colchicine treatment. The resistance seen in TifNV-High O/L may also come from TxAG-6 since smut resistance has been reported previously in a [(A. correntina x A. cardenasi) x A. batizocoi]x4 amphiploid (de Blas et al., 2016). Several spanish cultivars no longer in production demonstrated moderate or high resistance to peanut smut. Toalson (Simpson et al., 1979) and Comet (released in 1971 by J.S. Kirby, Oklahoma Agricultural Experiment Station) were resistant to T. frezzii, but both have only been evaluated for one growing season in our trials thus far. Toalson has previously been evaluated by others in Argentina and demonstrated smut resistance (Baldessari, personal communication). Toalson has an unusually thick hull, which may contribute to its resistance to the pathogen. OLin (Simpson et al., 2003) and Tamspan 90 (Smith et al., 1991) were both derived from crosses involving Toalson, and demonstrated moderate resistance at 5.5% (2 years) and 6.5% (1 year) disease incidence, respectively. Southwest Runner (Kirby et al., 1998), which resulted from a cross between Comet and Florunner (Norden et al., 1969), was also resistant to T. frezzii, exhibiting no disease during the one growing season it was evaluated. Toalson, Comet, and Southwest Runner also have a common ancestor in the cultivar Starr (Simpson, 1972b), which may be the potential source of their resistance. Starr was developed from a cross between Spantex (Simpson, 1972a) and PI 161317, a spanish-type line collected from Uruguay in 1947, that also appears in the pedigrees of six other spanish-type cultivars- Comet, Tamnut 74 (Simpson and Smith, 1975), Toalson, Pronto (Banks and Kirby, 1983), Spanco (Kirby et al., 1989), and Tamspan 90, according to Isleib and colleagues (2001).
To date, we have evaluated 208 peanut genotypes. comprised of germplasm accessions, breeding lines, and cultivars, for resistance to T. frezzii. Among those evaluated for three years, we identified eight entries that appear immune to smut, averaging 0% incidence. An additional eight entries have exhibited no infection by T. frezzii over a 2-year period. No association among collection origin, taxonomy, or growth habit was found among those accessions exhibiting complete resistance to smut. These data indicate that resistance to peanut smut exists in the U.S. peanut germplasm collection and can be incorporated into cultivars suitable for commercial production. While completely resistant PI accessions were found, none of the cultivars or breeding lines evaluated for three years in this study exhibited complete resistance. However, several did exhibit moderate resistance and could serve as potential parents for the development of resistant cultivars if crossed with immune PIs. Because the germplasm used in this study was purified by SSD and increased before evaluation, the entries identified as immune will serve as a valuable resource for peanut breeding programs across the globe.
Highly resistant and susceptible materials identified in these studies have been used to develop recombinant inbred line (RIL) populations for mapping molecular markers associated with peanut smut resistance. Traditional and experimentally aggressive approaches will be used in tandem to develop these mapping populations. The identified molecular markers will be deployed for use by peanut breeding programs and will allow rapid screening of breeding and other populations for peanut smut resistant materials, thus increasing the breeding efficiency and reducing the number of materials that must be phenotyped in the field.
Acknowledgements
Funding for this project was provided by USDA ARS CRIS Project 3072-21220-008-00D, the USDA ARS National Plant Disease Recovery System (NPDRS) and The MARS-Wrigley Company. The authors declare no conflict of interest. The authors would like to thank Lisa Myers, Sergio Morichetti (AGD), Guillermo de la Barrera and the personnel of INTA Manfredi’s Pathology Lab for technical assistance. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. USDA is an equal opportunity provider and employer.
Literature Cited
Arias R., Cazón L.I., Massa A.N., Scheffler B.E., Sobolev V.S., Lamb M.C., Duke M.V., Simpson S.A., Conforto C., Paredes J.A., Buteler M.I., and Rago A.M.. 2019. Mitogenome and nuclear-encoded fungicide target genes of Thecaphora frezii-causal agent of peanut smut. Fungal Genom. & Biol. 9: 160. https://digitalcommons.unl.edu/usdaarsfacpub/2198.
Banks D.J. and Kirby J.S.. 1983. Registration of ‘Pronto’ peanut. Crop Sci. 23:184.
Bressano M. Massa A.N., Arias R.S., de Blas F., Oddino C., Faustinelli P.C., Soave S., Soave J.H., Pérez M.A., Sobolev V.S., Lamb M.C., Balzarini M., Buteler M.I., and Seijo J.G.. 2019. Introgression of peanut smut resistance from landraces to elite peanut cultivars (Arachis hypogaea L.). PLoS One, 14. doi: [: 10.1371/journal.pone.0211920].
Carranza J. M., and Lindquist J.C.. 1962. Thecaphora frezii sp., parásita de Arachis sp. Boletín de la Sociedad Argentina de Botánica, 10:11–18.
Camiletti B.X., Paredes J.A., Monguillot J.H., Grosso N.R., and Rago A.M.. 2021. Fungicide efficacy of nanocrystal-based formulations against peanut smut. Crop Prot. 143:105522. doi: 10.1016/j.cropro.2020.105522Cazón, L.I. and A.M. Rago. 2016. Molecular detection of Thecaphora frezii in peanut (Arachis hypogaea L.) seeds. J. Plant Pathol. 98:327-330. doi:10.4454/JPP.V98I2.034.
Cazón LI, Paredes JA, Bisonard EM, and Rago A.M.. 2016. Capacidad de infección de Thecaphora frezii en el tiempo. 31° Jornada Nacional de Maní. 22 de septiembre. General Cabrera, Córdoba; p.52.
Cazón L.I., Paredes J.A., and Rago A.M.. 2018. The biology of Thecaphora frezii smut and its effects on Argentine peanut production. In: Adv. Plant Pathol. pp. 31-46. Kimatu, J.N., Editor. Intechopen. doi: 10.5772/INTECHOPEN.75837.
Chen C.Y., Barkley N.A., Wang M.L., Holbrook C.C., and Dang P.M.. 2014. Registration of purified accessions for the U.S. peanut mini-core germplasm collection. J. Plant Reg. 8:77–85.
Cignetti MI, Baldessari JMarraro Acuña F, and Mazzini P.H.. 2010. Evaluación multianual de cultivares de maní frente al carbón (Thecaphora frezii)). In: XXV Jornada Nacional del Maní, Córdoba, Argentina, 16 September 2010, 20-22.
de Blas F; Scheffler B; Arias R; Bressano M; Soave S; Pepermans M; Oddino C; Moresi A; Soave J; Buteler M. and Seijo G.. 2016. Evaluación del polimorfismo de un set de marcadores microsatélites en los progenitores de una población de RILs de maní segregante para enfermedades fúngicas. In: XXXI Jornada Nacional del Maní, pg 19-20.
Farías A., Baldessari J., Marraro A.F., and Mazzini P.H.. 2011. Evaluation of high-oleic peanut cultivars in the presence of peanut smut (Thecaphora frezii). In: 26th Jornada Nacional del Maní, General Cabrera, Córdoba, Argentina. pp. 20–22.
Food and Agriculture Organization of the United Nations, FAOSTAT. 2020. Available: http://faostat.fao.org
Ganuza M., Pastor N., Erazo J., Andrés J., Reynoso M.M., Rovera M., and Torres A.M.. 2017. Efficacy of the biocontrol agent Trichoderma harzianum ITEM 3636 against peanut smut, an emergent disease caused by Thecaphora frezii. Eur. J. Plant Pathol.151: 257-262. https://doi.org/10.1007/s10658-017-1360-0.
[GRIN] Germplasm Resources Information Network. 2021. U.S. Dept. of Agric. Nat. Plant Germ. Sys. Available at https://npgsweb.ars-grin.gov/gringlobal/crop.aspx?id¼86 (verified 30 March 2021).
Gorbet D.W. and Shokes F.M.. 2002. Registration of ‘C-99R’ peanut. Crop Sci. 42: 2207.
Holbrook C.C. and Dong W.. 2005. Development and evaluation of a mini-core collection for the U.S. peanut germplasm collection. Crop Sci. 45:1540-1544.
Holbrook C.C., Ozias-Akins P., Chu Y., Culbreath A.K., Kvien C.K., and Brenneman T.B.. 2017. Registration of ‘TifNV-High O/L’ peanut. J. Plant Reg. 11:229-230. doi: [: 10.3198/jpr2016.10.0059crc].
Isleib T.G., Holbrook C.C. and Gorbet D.W.. 2001. Use of plant introductions in peanut cultivar development. Peanut Sci. 28:96-113.
Kirby J.S., Banks D.J., and Sholar R.S.. 1989. Registration of ‘Spanco’ peanut. Crop Sci. 29:1573-1574.
Kirby J.S., Melouk H.A., Stevens T.E., Banks D.J., Sholar J.R., Damicone J.P., and Jackson K.E.. 1998. Registration of ‘Southwest Runner’ peanut. Crop Sci. 38: 546.
Marinelli A., March G., and Rago A.. 1995. El carbón del maní. Thecaphora frezii, sobre Arachis hypogaea L., Resúmenes VII Congreso de Micología y XVII Jornadas Argentinas de Micología. Rosario Santa Fe p. 134.
Marinelli A., March A., and Oddino C.. 2008. Aspectos biológicos y epidemiológicos del carbón del maní (Arachis hypogaea L.) causado por Thecaphora frezii Carranza & Lindquist. AgriScientia,, 25. doi: http://dx.doi.org/10.31047/1668.298x.v25.n1.2735.
Norden A.J., Lipscomb R.W., and Carver W.A.. 1969. Registration of ‘Florunner’ peanut. Crop Sci. 9: 850.
Oddino C., Soave J., Soave S., Moresi A., Bianco C., Buteler M., Faustinelli P., and Torre D.. 2013. Genetic advances in the tolerance of peanut to smut caused by Thecaphora frezzii.. In 288 Jornada Nacional del Maní, General Cabrera, Córdoba, Argentina. pp. 28–30.
Oddino C.M., Soave J.A., Soave S.J., Buteler M.I., Moresi A., deBlas F.J. 2017. Sources of smut resistance in peanut wild species and Bolivian landraces. In: Advances in Arachis through genomics and biotechnology. Cordóba, Argentina. 14-17 March 2017. pp. 8.
Paredes J.A. Cazón L.I. Oddino C., Monquillot J.H., Rago A.M., and Molina J.P.E.. 2021. Efficacy of fungicides against peanut smut in Argentina. Crop Prot. 140:105403. doi: [: 10.1016/j.cropro.2020.105403].
RStudio Team. 2020. RStudio: Integrated Development for R. RStudio, PBC, Boston, MA URL http://www.rstudio.com/.
Rago A.M., Cazón J.A., Molina J.P.E., and Conforto E.C.. 2017. Peanut Smut: From an emerging disease to an actual threat to Argentine peanut production. Plant Dis. 101:400-408.
SAS Institute Inc. 2013. SAS/ACCESS® 9.4 Interface to ADABAS: Reference. Cary, NC: SAS Institute Inc.
Saxton A.M. 2000. A Macro for Converting Mean Separation Output to Letter Groupings in PROC MIXED. http://www.math.wsu.edu/faculty/djohnson/resources/STAT512/SAS%20Material/pdmix800.sas (Accessed 7/2021).
Simpson C.E. 1972a. Registration of ‘Spantex’ peanut. Crop Sci. 12:395.
Simpson C.E. 1972b. Registration of ‘Starr’ peanut. Crop Sci. 12:395.
Simpson C.E., Baring M.R., Schubert A.M., Melouk H.A., Lopez Y., and Kirby J.S.. 2003. Registration of ‘OLin’ peanut. Crop Sci. 43:1880-1881.
Simpson C.E. and Smith O.D.. 1975. Registration of ‘Tamnut 74’ peanut. Crop Sci. 15:603-604.
Simpson C.E., Smith O.D., and Boswell T.E.. 1979. Registration of ‘Toalson’ peanut. Crop Sci. 19:742.
Smith O.D., Simpson C.E., Grichar W.J., and Melouk H.A.. 1991. Registration of ‘Tamspan 90’ peanut. Crop Sci. 31: 1711.
Soave J.A., Bianco C., Burgoa R., Montaño R., Rago A.M., and Cazón L.I.. 2014. First detection of peanut smut (Thecaphora frezii) in Bolivia. In: Ploper L.D. (ed.) Proceedings of the 38 Congreso Argentino de Fitopatologı´ a, San Miguel de Tucuma´ n, Argentina. 4-6 June 2014. Asoc. Argentina Fitopatol., Córdoba, Argentina. pp. 211.
Wann D.Q., Falco A., Cavigliasso M., and Cassano C.. 2020. Phenotypic variation of peanut smut (Thecaphora frezii) incidence and severity in the U.S. peanut mini-core collection. Peanut Sci. 47:46-53.
Notes
- USDA ARS Wheat, Peanut, and Other Field Crops Research Unit, Stillwater, OK 74075 [^]
- Instituto Nacional Tecnología Agropecuaria (INTA), Manfredi, Argentina [^]
- Hudson Alpha Institute for Biotechnology, 601 Genome Way Northwest, Huntsville, AL 35806 [^]
- USDA ARS, Crop Genetics and Breeding Research Unit, Tifton, GA 31793 [^]
- USDA-ARS, Plant Genetic Resources Conservation Unit, Griffin, GA 30223 [^]
- University of Georgia, Institute of Plant Breeding, Genetics, and Genomics, Tifton, GA 31793 [^]
- Instituto Nacional Tecnología Agropecuaria (INTA), Marcos Juarez, Argentina [^]
- Department of Biomedical Sciences, Rocky Vista University, Parker, CO, 80134 [^] Corresponding Author: Kelly D. Chamberlin , USDA ARS Wheat, Peanut, and Other Field Crops Research Unit1301 N. Western Rd., Stillwater, OK 74075; kelly.chamberlin@usda.gov; 405-385-5432