

Introduction
Spotted wilt is a systemic disease in peanut (Arachis hypogaea L.) caused by Tomato spotted wilt virus (TSWV, genus Tospovirus, family Bunyaviridae) (Moyer, 1999). TSWV is one of the most widely distributed and economically important plant viruses with a wide host range infecting more than 800 species of both monocotyledonous and dicotyledonous plants (Prins and Goldbach, 1998). In the southeastern United States, TSWV is a serious problem in peanut with estimated economic losses in excess of $40 million (Brown et al., 2005). It also causes significant crop losses in pepper, tobacco and tomato (Padgett et al., 1995) and can infect weeds (Chatzivassiliou et al., 2007) that may act as potential over-wintering reservoirs of the virus. TSWV is efficiently transmitted by several thrips species (Whitfield et al., 2005) including the western flower thrips, Frankliniella occidentalis (Sakimura, 1962), and the tobacco thrips, F. fusca (Hinds) (Sakimura, 1963), both of which are present in most of the peanut growing areas of the United States (Culbreath et al., 2003). Only thrips that acquired TSWV in the larval stage and adults derived from such larvae are able to transmit the virus (Nagata et al., 2002). Viruliferous adults can continue to feed and infect peanut plants for the remainder of their entire adult lives (Wijkamp et al., 1993). Peanut infection by TSWV can produce a wide range of symptoms including concentric ring spots, leaf chlorosis, stunting, deformity and discoloration of pods and kernels (Culbreath et al., 1992).
Current standard determination of TSWV intensity in the field relies on a visual disease rating that represents both disease incidence and severity (Wells et al., 2002; Cantonwine et al., 2006). However, it has been determined that asymptomatic plants can still be TSWV-infected (Culbreath et al., 1992a; Rowland et al., 2005). Assays such as enzyme-linked immunosorbent assay (ELISA) or reverse transcription polymerase chain reaction (RT-PCR) have been utilized to detect TSVW in peanut (Jain et al., 1998; Rowland et al., 2005; Do Nascimento et al., 2006; Murakami et al., 2006; Wang et al., 2007). The ELISA technique is based on the ability of antibodies to recognize proteins, typically coat protein, from a specific virus of interest. The level of viral proteins present at any one time in a specific tissue is a dynamic relationship between protein synthesis versus protein degradation by the plant's defense system (Delaure et al., 2008). The RT-PCR assay utilizes viral RNAs in various tissues and converts them into cDNAs by reverse transcription (RT). Specific primers are then utilized in a polymerase chain reaction (PCR) and infected or non-infected individuals are scored based on the presence or absence of PCR product bands in agarose gel electrophoresis. Comparisons between ELISA and RT-PCR assays to detect TSWV in field-grown chrysanthemum have been reported (Matsuura et al., 2002; Matsuura et al., 2004). To date, there are no reports comparing ELISA and RT-PCR in detecting TSWV in peanut tissues. The goal of this study was to compare the efficacy of ELISA and RT-PCR in determining TSWV infection on field-grown peanut and evaluate their use as a diagnostic tool to study TSWV epidemiology in peanut.
Materials and Methods
Field collection of peanut roots
The efficacy of ELISA and RT-PCR for detection of TSWV was compared in two separate experiments. In both experiments, peanut root crown samples were used for analysis based on reports of higher incidence of TSWV in root crown compared to foliar tissue in different peanut cultivars (Rowland et al., 2005; Murakami et al., 2006). For both experiments, plants were randomly selected and roots were cut just above the root crown, put into plastic bags and placed on ice until returning to the laboratory for further processing. TSWV positive controls were obtained from symptomatic field-grown peanut plants located at the National Peanut Research Laboratory, Dawson, GA. Infection was confirmed using ImmunoStrips (Agdia, Elkhart, IN) and by RT-PCR.
Experiment 1
Peanut root samples were collected at the Bolton Farm (Terrell County, GA). The crop was planted on 19 April 2006. In 2007, the crop was planted on two dates: 20 April and 22 May. Three peanut cultivars, Georgia Green (Branch, 1996), Georgia-02C (Branch, 2003), and AP-3 (Gorbet, 2007), were planted at all dates. In this experiment, root samples were split vertically into two equal halves. One half of the root crown was used for ELISA analysis while the other half was stored at −20 C for RNA extraction. Commercially available TSWV-specific Double-antibody sandwich enzyme-linked immunosorbant (DAS-ELISA) kit and ImmunoStrips (Agdia Inc., Elkhart, IN, USA) were used. For DAS-ELISA, a 100 µl of 1/200 dilution of anti-TSWV coating antibody was placed into each well of a 96-well ELISA plate and left at 4 C overnight, then washed three times with washing buffer. Dried root samples (approximately 0.2 g) were ground in 2 ml of sample extraction buffer, and from each sample 100 µl was loaded into a well of the pre-coated plate and incubate at 4 C overnight. The plate was then washed three times with washing buffer and 100 µl aliquots of 1/200 diluted anti-TSWV alkaline phosphatase enzyme conjugate was added to each well and incubate at room temperature for 2 hours. The plate was washed as described above and 100 µl aliquots of Para-nitro phenol phosphate (PNP) solution at 1 mg/mL was added to each well and incubate for 60 minutes at room temperature. Absorbance measurements were performed using a microplate reader (Bio Tek, Winooski, VT). Absorbance readings for two negative control wells were averaged, multiplied by three and added to twice the standard deviation of the negative controls. This value was used as a base line. Absorbance values above the base line were considered positive readings for the presence of TSWV. For the ImmunoStrip assay 0.15 g of tissue was weighed out and placed into a sample bag preloaded with 3 ml of sample extraction buffer. Samples were pulverized with a plastic lined mallet to a fine consistency. ImmunoStrips were placed into sample bags to the marked line and reactions were allowed to proceed (from a minimum of 5 minutes to a maximum time of 30 minutes). Strips were scored as positive (dark test line and dark control line) and negative (no test line and dark control line) and recorded.
For RT-PCR analysis, root samples (∼100 mg) were ground to a fine powder in liquid nitrogen using a pre-chilled mortar and pestle. Total RNA was extracted and DNased using RNeasy Plant Mini Kit (Qiagen, Valencia, CA) according to the manufacturer's instruction. RNA concentrations were measured using a spectrophotometer (Thermo Fisher, Waltham, MA) and the quality of RNA was determined by RNA gel electrophoresis. After RNA quality analysis was performed, first strand cDNA was synthesized using 0.75 µg total RNA. Total RNA was denatured at 95 C for 5 minutes in the presence of TSWV 722 and 723 primers (50 µM each) (Adkins and Rosskopf, 2002) in a total volume of 14 µl and allowed to anneal for 10 minutes at 45 C. An aliquot of 36 µl of reaction mixture (3 µl RNase-Free water, 10 µl 5X MMLV RT buffer, 16 µl 2 mM dNTP mix, 4 µl 100 mM DTT, 2 µl MMLV-RT [200 u/µl], 1 µl of RNAsin [40 u/µl]) was added. The RT reactions were performed at 45 C for 45 minutes, followed by 5 minutes at 95 C, 5 minutes at 22 C, and cooled to 4 C. One microliter of RT reaction was used as a template for the PCR reaction. Twenty-four microliters of the PCR reaction mixture (10 µl sterile water, 12.5 µl Green GOTAQ mix [Promega, Madison, WI], 0.75 µl TSWV 722 primer [100 µM] and 0.75 µl TSWV 723 [100 µM] primers) were added. The PCR profile consisted of a denaturing step at 95 C for 2 minutes, followed by 30 cycles of 95 C for 45 seconds, 55 C for 45 seconds, 72 C for 1 minute, followed by a 72 C extension for 10 minutes, and a final hold step at 4 C. PCR products were separated on a 1.5% TAE agarose gel. Gel image was captured on Gel Logic 200 Imaging System (Kodak, Rochester, NY) and scored visually for the presence or absence of PCR bands.
Experiment 2
Peanut (cv. Georgia Green) root samples were collected in a grower field in Bulloch County, GA. The peanut crop was planted on the week of 22 May 2006. In this experiment, a long-term storage technique was utilized in which roots were cut horizontally into slices and allowed to dry in weigh boats at room temperature for two to three days. Dried root slices were placed into 15 ml storage tubes preloaded with Drierite Absorbent (Fisher, Pittsburg, PA) and stored at −20 C. DAS-ELISA was performed as described above. RT-PCR analysis was identical as that used in Experiment 1 except using the dried samples for analysis.
Statistical Analysis
Comparison of TSWV infection rates as detected by RT-PCR and ELISA were analyzed using PROC FREQ procedure in SAS version 9.1 (SAS Institute, Cary, North Carolina) with the CHISQ TESTP option in the Model statement utilizing chi-square goodness of fit test. The ELISA observed results were used as the standard for expected values.
Results
Experiment 1
In 2006 at the Bolton Farm, 11 out of 24 total samples (45.8%) tested positive for TSWV by RT-PCR and 9 (37.5%) by ELISA (Table 1). Twenty samples (83.3%) showed identical results in both assays (8 positives and 12 negatives), while three samples (12.5%) tested TSWV positive by RT-PCR but negative by ELISA and 1 sample (4.2%) tested TSWV negative by RT-PCR but positive by ELISA. No statistical difference was observed between ELISA and RT-PCR results (χ2 = 0.625, P> 0.4290) when Bolton 2006 data was analyzed together.
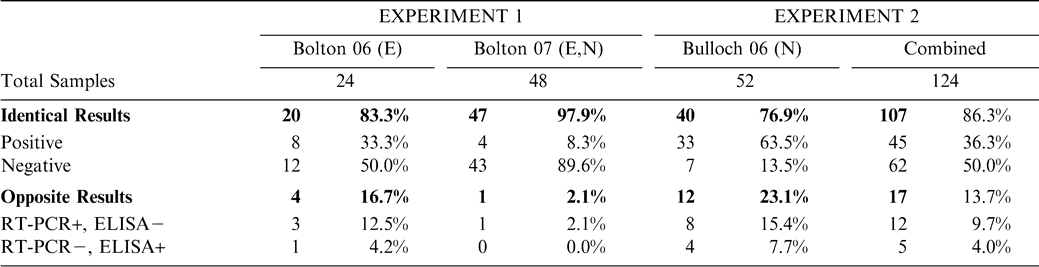
Comparison of RT-PCR and ELISA assays for TSWV detection in peanut root tissues. Results are shown for total samples collected in Experiment 1, Bolton 2006 (E = early planting) and Bolton 2007 (E and N = normal planting); Experiment 2, Bulloch 2006 (N) and all samples combined. Experiment 1 used fresh root tissue and experiment 2 used dried, frozen root tissue.
From both planting dates at Bolton in 2007, a total of 48 samples were collected and tested. A total of 5 samples (10.4%) tested TSWV positive by RT-PCR and 4 (8.3%) tested positive by ELISA (Table 1). Further analysis showed 47 (97.9%) out of 48 samples showed identical test results (4 positives and 43 negatives) for both assays. Only 1 sample (2.1%) tested TSWV positive by RT-PCR but was negative by ELISA. No statistical difference was also observed comparing ELISA and RT-PCR results (χ2 = 0.381, P > 0.5370) when Bolton 2007 data was analyzed together. Because there were no observed statistical differences in either Bolton 2006 or 2007 data set, results were pooled and statistically analyzed. Again, no statistical difference was observed between ELISA and RT-PCR results (χ2 = 0.869, P > 0.3510) when all 72 samples were analyzed together.
Experiment 2
In 2006 at the Bulloch County, GA site, a total of 52 samples were tested. Forty-one samples (78.8%) tested TSWV positive by RT-PCR and 37 (71.2%) tested positive by ELISA (Table 1). Among the 52 samples analyzed, 40 samples (76.9%) showed identical results (33 positives and 7 negatives) for both assays. Eight samples (15.4%) tested positive by PCR but tested negative by ELISA; while 4 samples (7.7%) tested negative by RT-PCR but positive by ELISA. There was no statistical difference comparing ELISA and RT-PCR results (χ2 = 1.555, P > 0.2120) when all Bulloch 2006 samples were analyzed together. When combining the TSWV testing results from both experiments 1 and 2, a total of 124 samples were collected and analyzed with both assays. A total of 57 samples (46.0%) were classified as positive for TSWV infection by RT-PCR and 50 samples (40.3%) by ELISA (Table 1). Among these 124 samples, 107 (86.3%) showed identical results (45 positives and 62 negatives) for both assays. Twelve samples (9.7%) tested TSWV positive by RT-PCR but tested negative by ELISA; 5 samples (4.0%) tested TSWV negative by RT-PCR but tested positive by ELISA. Statistical analysis utilizing chi-square goodness of fit test with ELISA results as a standard (χ2 = 1.840, P > 0.1750) showed no significant statistical difference between ELISA and RT-PCR results when all data were analyzed together.
Discussion
ELISA has become the standard assay for TSWV diagnosis in peanut (Rowland et al., 2005; Murakami et al., 2006; Wang et al., 2007). The development of RT-PCR for TSWV diagnosis (Adkins and Rosskopf, 2002; Dietzgen et al., 2005; Jain et al., 1998; Mumford et al., 1996) provides another assay to detect TSWV in peanut. Our goal is to assess the two methods to determine if results are comparable. Root crown tissues were selected since there have been reports of higher incidences of TSWV in root crowns compared to leaves (Rowland et al., 2005; Murakami et al., 2006). In this study, 124 root samples were collected from field grown peanut plants and subjected to both ELISA and RT-PCR assays and the results were compared. While only a small number of plants were analyzed both ELISA and RT-PCR were able to detect the same infected plants that exhibited no viral symptoms. No statistical difference was observed which indicates that ELISA and RT-PCR are comparable methods for diagnosis of TSWV incidence in field-grown peanut.
The results reported here are similar to previous reports comparing ELISA and RT-PCR for detection of Prunus necrotic ring spot virus and Prune dwarf virus in almond (Mekuria et al., 2003). In a separate study comparing the efficacy of ELISA and RT-PCR in detecting TSWV, results showed that 3 asymptomatic, infected chrysanthemum plants (out of 50 plants) were identified to be infected by both assays (Matsuura et al., 2002). However, some studies have found more marked differences between ELISA and RT-PCR for other viruses. For example, a study in apples evaluating the detection levels of Apple mosaic virus, Apple stem grooving virus, and Apple chlorotic leaf spot virus using ELISA showed a 36% infection level compared to a 44% using RT-PCR (Caglayan et al., 2006).
The quality of RNA was evaluated for the extraction from fresh root tissues (experiment 1) and dried, frozen root tissues (experiment 2). The result indicates that both types of tissue can be efficiently processed for both ELISA and RT-PCR assays without loss to the relative quality of viral proteins and viral RNA, respectively. The ability of dried, frozen roots to maintain testing consistency would eliminate the need to specially treat samples prior to processing and reduce the need for −80 C freezer space.
Serological kits are commercially available and fairly economical. Two formats are currently available, ImmunoStrips or DAS-ELISA (Agdia). The ImmunoStrips are easy to perform, can be completed in the field within minutes, do not require lab equipment, and the cost per sample is reasonable. DAS-ELISA requires minimal laboratory facilities and an operator with laboratory experience, but is more economical for large number of samples. On the other hand the RT-PCR assay is estimated to cost over 20 times more than ELISA and requires expensive equipment and an operator with a high level of laboratory experience. The advantage of this assay is that the purified RNA samples can be further processed using real-time RT-PCR to actually quantify viral RNA in specific tissues. However, because of the relative ease and inexpensive cost of ELISA versus RT-PCR, and the results of the current study showing no difference in detection of TSWV, ELISA remains the best choice for routine evaluation of TSWV infection in peanut.
Acknowledgements
We thank Larry Powell and Kathy Gray for technical assistance in the field and in the laboratory. We thank Ben Mullinix for statistical analysis and consultation.
Literature Cited
Adkins S. and Rosskopf E. N. 2002 Key West nightshade, a new experimental host for plant viruses. Plant Dis 86 : 1310 – 1314 .
Branch W. D. 1996 Registration of ‘Georgia Green’ Peanut. Crop Sci 1996 36 : 806 .
Branch W. D. 2003 Registration of ‘Georgia-02C’ Peanut. Crop Sci 2003 43 : 1883 – 1884 .
Brown S. L. , Culbreath A. K. , Todd J. W. , Gorbet D. W. , Baldwin J. A. , and Beasley J. P. 2005 Development of a method of risk assessment to facilitate integrated management of spotted wilt in peanut. Plant Dis 89 : 348 – 356 .
Caglayan K. , Ulubas Serce C. , Gazel M. , and Jelkmann W. 2006 Detection of four apple viruses by ELISA and RT-PCR assays in Turkey. Turk J. Agric. For 30 : 241 – 246 .
Cantonwine E. G. , Culbreath A. K. , Stevenson K. L. , Kemerait R. C. , Brenneman T. B. , Smith N. B. , and Mullinix B. G. 2006 Integrated disease management of leaf spot and spotted wilt of peanut. Plant Dis 90 : 493 – 500 .
Chatzivassiliou E. K. , Peters D. , and Katis N. I. 2007 The role of weeds in the spread of Tomato spotted wilt virus by Thrips tabaci (Thysanoptera: Thripidae) in tobacco crops. J. Phytopathol 155 : 699 – 705 .
Culbreath A. K. , Todd J. W. , and Demski J. W. 1992 Production of Florunner peanut infected with Tomato spotted wilt virus. Peanut Sci 19 : 11 – 14 .
Culbreath A. K. , Todd J. W. , and Brown S. L. 2003 Epidemiology and management of Tomato spotted wilt in peanut. Annu. Rev. Phytopathol 41 : 53 – 75 .
Delaure S. L. , Hemelrujck W. V. , De Bolle M. F. C. , Cammue B. P. A. , and De Coninck B. M. A. 2008 Building up plant defenses by breaking down proteins. Plant Sci 174 : 375 – 385 .
Dietzgen R. G. , Twin J. , Talty J. , Selladurai S. , Carroll M. L. , Coutts B. A. , Berryman D. I. , and Jones R. A. C. 2005 Genetic variability of Tomato spotted wilt virus in Australia and validation of real time RT-PCR for its detection in single and bulked leaf samples. Ann. App. Biol 146 : 517 – 530 .
Do Nascimento L. C. , Pensuk V. , Da Costa N. P. , De Assis Filho F. M. , Pio-Ribeiro G. , Deom C. M. , and Sherwood J. 2006 Evaluation of peanut genotypes for resistance to Tomato spotted wilt virus by mechanical and thrips inoculation. Pesq. Agropec. Bras 41 : 937 – 942 .
Gorbet D. W. 2007 Registration of ‘AP-3’ Peanut. J. of Plant Registrations 1 : 126 – 127 .
Jain R. K. , Pappu S. S. , Pappu H. R. , Culbreath A. K. , and Todd J. W. 1998 Molecular diagnosis of Tomato spotted wilt tospovirus infection of peanut and other field and greenhouse crops. Plant Dis 82 : 900 – 904 .
Matsuura S. , Hoshino S. , Yayashi H. , Kohguchi T. , Hagiwara K. , and Omura T. 2002 Effects of latent infection of stock plants and abundance of thrips on the occurrence of Tomato spotted wilt virus in Chrysanthemum fields. J. Gen. Plant Pathol 68 : 99 – 102 .
Matsuura S. , Ishikura S. , Shigemoto N. , Kajihara S. , and Hagiwara K. 2004 Localization of Tomato spotted wilt virus in Chrysanthemum stock plants and efficiency of viral transmission from infected stock plants to cuttings. J. Phytopathol 152 : 219 – 223 .
Mekuria G. , Ramesh S. A. , Alberts E. , Bertozzi T. , Wirthensohn M. , Collins G. , and Sedgley M. 2003 Comparison of ELISA and RT-PCR for the detection of Prunus necrotic ring spot virus and prune dwarf virus in almond (Prunus dulcis). J. Virol. Meth 114 : 65 – 69 .
Moyer J. W.
1999
Tospoviruses (Bunyaviridae).
1803
–
1807
In
Granoff A.
and
Webster R. G.
Encyclopedia of Virology
Academic Press
San Diego, CA
.
Mumford R. A. , Barker I. , and Wood K. R. 1996 An improved method for the detection of Tospoviruses using the polymerase chain reaction. J. Virol. Meth 57 : 109 – 115 .
Murakami M. , Gallo-Meagher M. , Gorbet D. W. , and Meagher R. L. 2006 Utilizing immunoassays to determine systemic tomato spotted wilt infection for elucidating field resistance in peanut. Crop Prot 25 : 235 – 243 .
Nagata T. , Inoue-Nagata A. K. , Lent J. V. , Goldbach R. , and Peters D. 2002 Factors determining vector competence and specificity for transmission of Tomato spotted wilt virus. J. Gen. Virol 83 : 663 – 671 .
Padgett G. B. , Bertrand P. F. , and Gay J. D. 1995 Spotted wilt in Georgia's crops. Univ. Georgia Coop. Ext. Serv. Bull 1127 .
Prins M. and Goldbach R. 1998 The emerging problem of tospovirus infection and nonconventional methods of control. Trends Microbiol 6 : 31 – 35 .
Rowland D. , Dorner J. , Sorensen R. , Beasley J. P. , and Todd J. 2005 Tomato spotted wilt virus in peanut tissue types and physiological effects related to disease incidence and severity. Plant Pathol 54 : 431 – 440 .
Sakimura K. 1962 Frankliniella occidentalis (Thysanoptera: Thripidae), a vector of the Tomato spotted wilt virus, with special references to the color forms. Ann. Entomol. Soc. Am 55 : 387 – 89 .
Sakimura K. 1963 Frankliniella fusca, an additional vector for the Tomato spotted wilt virus, with notes on Thrips tabaci, another vector. J. Phytopathol 53 : 412 – 15 .
Wang M. L. , Pinnow D. L. , and Pittman R. N. 2007 Preliminary screening of peanut germplasm in the U.S. collection for TSWV resistance and high flavonoid content. Plant Pathol. J 6 : 219 – 226 .
Wells M. L. , Culbreath A. K. , Todd J. W. , Brown S. L. , and Gorbet D. W. 2002 A regression approach for comparing field resistance of peanut cultivars to tomato spotted wilt tospovirus. Crop Prot 21 : 467 – 474 .
Whitfield A. E. , Ullman D. E. , and German T. L. 2005 Tospovirus-thrips interactions. Annu. Rev. Phytopathol 43 : 459 – 489 .
Wijkamp I. , Van Lent J. , Kormelink R. , Goldbach R. , and Peters D. 1993 Multiplication of tomato spotted wilt virus in its insect vector, Frankliniella occidentalis. J. Gen. Virol 74 : 341 – 349 .
Notes
1Biochemist, Plant Physiologist, and Research Agronomist, USDA-ARS, National Peanut Research Laboratory, PO Box 509, 1011 Forrester Dr. SE, Dawson, GA 39842. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the USDA.
*Corresponding author (email: Phat.Dang@ars.usda.gov).