

Introduction
Past research has shown differential crop cultivar response to herbicides (Brecke, 1989; Hardcastle, 1974; Hauser et al., 1981; Johnson et al., 1992, 1993; Knauft et al., 1990; Smith and Wilkinson, 1974). Sweet corn (Zea mays L.) cultivars responded differently to chloroacetamide and thiocarbamate herbicides (Bennett and Gorski, 1989). Soybean [Glycine max L. (Merr.)] cultivars have shown a differential response to metribuzin (Hardcastle, 1974; Smith and Wilkinson, 1974) and imazapic (experimental code AC 263,222) (Wixson and Shaw, 1991). Peanut cultivars have also shown a differing response to imazapic (Richburg et al., 1994).
Hauser et al., (1976) reported that Goldkist GK 3 was sensitive to multiple applications of dinoseb in two of eight trials. However, in later research with six peanut cultivars, no negative effects were observed, and the sensitivity of Goldkist GK 3 was attributed to weed competition (Hauser et al., 1981). Recent studies have evaluated the response of peanut cultivars to herbicides (Brecke, 1989; Johnson et al., 1992, 1993; Knauft et al., 1990; Wehtje et al., 1991). Brecke (1989) reported greater sensitivity of Early Bunch and Southern Runner to paraquat compared to Florunner. However, Wehtje et al., (1991) reported that Florunner, NC 7, Southern Runner, and Sunrunner did not exhibit differential sensitivity to a single application at 0.14 kg ai/ha. Later research by Knauft et al., (12) reported that two paraquat applications at 0.14 kg/ha reduced yields of Florunner, Southern Runner, and Sunrunner. Peanut cultivar response has also been reported with chlorimuron as well (Brecke, 1989; Johnson et al., 1992). Johnson et al., (1993) reported there was no yield response of Florunner, NC 7, or Southern Runner to multiple herbicide applications, but canopy widths were affected. Diclosulam, an ALS-inhibiting herbicide like imazethapyr and imazapic did not affect growth, disease incidences, grade parameters, or yield in eight virginia peanut cultivars (Bailey et al., 2000).
Imazethapyr is registered for use in peanut as a PPI, PRE, EPOST, or POST treatment. Imazethapyr provides soil residual control of numerous broadleaf weeds and perennial sedges (Richburg et al., 1995, 1996; Wilcut et al., 1991a, 1991b, 1994). The registration of new peanut herbicides necessitates determination if tolerance differs among peanut cultivars (Hammons, 1976). Since the 1970's, Florunner has been the predominant cultivar. However, recent hectarage increases in other runner cultivars and production of virginia-type cultivars in regions previously dominated by runners has occurred (Holbrook and Kvien, 1989). In addition, spanish cultivars are grown extensively in the Texas-Oklahoma production area (Hammons, 1976).
The objectives of this research were to evaluate the response of eight peanut cultivars in Georgia and five in Texas to EPOST applications of imazethapyr.
Materials and Methods
Experiments were conducted in 1992 and 1993 at the Coastal Plain Experiment Station located near Tifton, GA and in Yoakum, TX. The soil type at Tifton was a Tifton loamy sand (fine-loamy, siliceous, thermic Kandiudults) with 0.5% organic matter and pH 6.2. The soil type at Yoakum was a Tremona loamy fine sand (thermic Aquic Arenic Palenstalfs) with 1% organic matter and pH 7.0. The experiments were kept weed free using pendimethalin applied at 1.1 kg ai/ha PPI for annual grass and small-seeded broadleaf weed control (Wilcut et al., 1994) and weekly hand weedings.
Plots were two rows wide spaced 90 cm apart by 6.1 m. Planting dates were May 11, 1992 and April 19, 1993 for Tifton and June 8, 1992 and June 7, 1993 for Yoakum. Peanuts were planted 5 cm deep, spaced 5, 8, and 5 cm apart for runner, virginia, and spanish peanut, respectively.
The experimental design was a randomized complete block at Tifton with a factorial arrangement of treatments with four replications in 1992 and three replications in 1993. The two herbicide treatments were either none or imazethapyr applied EPOST at 71 g ai/ha plus a nonionic surfactant (X-77) at 0.25% v/v of the spray volume. The eight peanut cultivars consisted of five runner- and three virginia-types. The runner-types were Florunner, Georgia Runner, Goldkist GK 7, Southern Runner, and Sunrunner and the virginia-types were NC 7, NC-V11, and AgraTech VC 1.
The experimental design at Yoakum was a randomized complete block with a factorial arrangement of treatments with four replications in 1992 and three replications in 1993. The five peanut cultivars consisted of one runner and four spanish-types. The runner-type was Florunner and the spanish-types were Pronto, Spanco, Starr, and Tamspan 90.
Imazethapyr was applied EPOST at 71 g/ha with a CO2 backpack sprayer and a nonionic surfactant (X-77) was included at 0.25% v/v of the spray volume. Chlorothalonil was applied every 10 to 14 days at 1.25 kg ai/ha to the experimental test areas for early (Cercospora arachidicola Hori.) and late [Cercosporidium personatum (Berk. & Curt.) Deighton] leafspot control, beginning 30 days after planting (DAP) and going till harvest at all locations. Harvest dates were determined using the Hull Scrape method on nontreated peanut of each cultivar grown in front of the experimental areas (Johnson, 1987). Runner- and virginia-type peanut were dug approximately 140 DAP except Southern Runner which was dug approximately 150 DAP. Southern Runner requires approximately 10 to 20 additional days to mature compared to most other cultivars (Richburg et al., 1994). Spanish-type peanut were dug approximately 130 DAP.
Canopy width measurements were taken at Tifton in 1992 at 4 and 6 weeks after treatment WAT, and 2, 4, 6, 8, and 10 WAT in 1993. Peanut yield and grade analysis were taken for all cultivars evaluated. A random 500-g sample free of foreign matter and splits due to harvesting, was taken for grade analysis. Grade analysis included the percentage total sound mature kernels (TSMK) (sound mature kernels plus sound splits), extra large kernels (ELK) for virginia-type peanut or jumbo for runner-type peanut (seed that rode a 8.33- by 25.40-mm slotted screen), medium kernels (seed that rode a 7.14- by 25.40-mm slotted screen for virginia-types and 7.14- by 19.0-mm for runner-types), and number 1 (No. 1) kernels (seed that rode a 5.95- by 25.40-mm for virginia-types and 6.36- by 19.0-mm for runner-types), other kernels (seed that passed through screens that retained No. 1 seed), and damaged kernels (visually damaged seed that passed through screens that retained No. 1 seed). Percentage fancy pods (unshelled peanut that rode a 13.5-mm roller spacing on the presizer) for virginia-types also was taken.
Evaluations for possible disease interactions with imazethapyr were also taken prior to harvest. Disease intensity of early leafspot and late leafspot was rated immediately prior to harvest. The Florida 1–10 rating scale was used where 1 = no spots and 10 = complete defoliation by leafspot, resulting in death (Chiteka et al., 1988). A visual estimate of plants with symptoms of spotted wilt (TSW) also was taken at this time. Immediately after peanut plants were inverted, the number of southern stem rot (Sclerotium rolfsii Sacc.) loci was determined for each plot, where a locus represented 31 cm or less of linear row with one or more plants diseased (Rodriquez-Kabana et al., 1975). These ratings were subsequently converted to a percentage of 31 cm sections of linear row with a least one diseased plant.
Canopy width and height were taken in 1992 in Yoakum at 3 and 6 WAT. Peanut yield and grade analysis were taken in 1992 and 1993. Grade analysis included percentage TSMK, other kernels, and damaged kernels. Grade analysis was conducted on a 200 g random sample from each plot. Screen sizes for grade analysis were as described previously for runner- and spanish-type peanut, with the exception of the No. 1 seed screen size for spanish-type peanut which was 5.95- by 19.05-mm.
Due to different soil type, cultivars, data taken, and geographical regions, experiments from Tifton and Yoakum were analyzed separately. Peanut yield and grade data, disease incidence, and canopy width measurements from 1992 at Tifton were subjected to analysis of variance, which revealed nonsignificant treatment-by-year interactions, consequently years were pooled. Canopy width measurements in 1993 at Tifton were subjected to regression analysis. In addition, analysis of variance was performed on regression coefficients of each regression equation at Tifton using individual replication data. Canopy height and width, peanut yield, TSMK, damaged and other kernels at Yoakum were subjected to analyses of variance that revealed non-significant treatment by year interactions, consequently years were pooled. Means from Tifton and Yoakum were separated with the appropriate Fisher's Protected Least Significant Difference (LSD) Test at the 5% level of probability.
Results and Discussion
Disease Interactions-tifton, Ga
The occurrence of early and late leafspot, TSW, and southern stem rot was independent of cultivar, imazethapyr, and the interactions thereof (data not shown).
Peanut Injury-tifton, Ga
Early season peanut injury in the form of minor stunting and chlorosis was less than 11% early-season. By mid-season, visual injury was not apparent (data not shown).
Peanut Canopy Width-tifton, Ga
There was not a significant cultivar by imazethapyr interaction on canopy width in 1992 at 4 or 6 WAT (Table 1). However, there was a significant cultivar main effect on canopy width in 1992 at 4 and 6 WAT. Canopy width differences between cultivars were expected since Florunner, Georgia Runner, Southern Runner, and Sunrunner have runner growth habits, while NC 7, NC-V11, and AgraTech VC 1 have decumbent growth habits (Richburg et al., 1994).
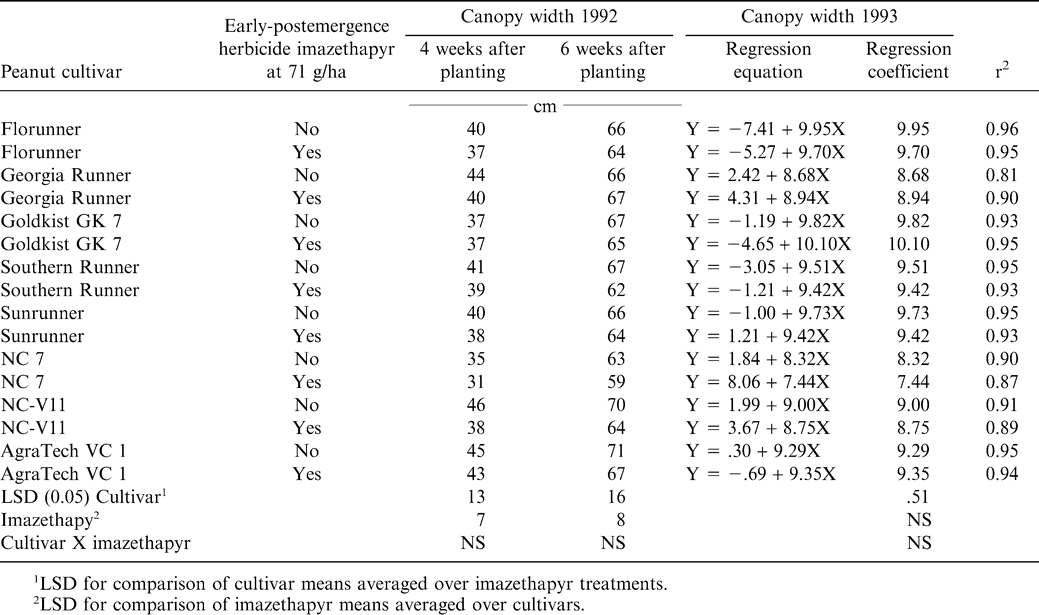
The main effect of imazethapyr on canopy width in 1992 at 4 and 6 WAT also was significant. Although the interaction of cultivar by imazethapyr was not significant, the canopy width of all cultivars treated with imazethapyr was numerically equal to or less than each respective nontreated cultivar in 1992, at 4 and 6 WAT. Previous research showed that peanut cultivars treated with imazapic also had smaller canopy widths 4 and 6 WAT (Richburg et al., 1994). One paraquat application at 0.14 kg/ha reduced canopy width of Florunner, Sunrunner, Georgia Runner, and NC 7 (Knauft et al., 1990). Johnson et al., (1993) reported two paraquat plus bentazon applications reduced canopy width of Florunner, Southern Runner, and NC 7.
Regression analysis of canopy width measurements in 1993 revealed a linear canopy width response on all cultivars (Table 1). Analysis of regression coefficients revealed a significant cultivar main effect. Regression coefficients of canopy width were used to indicate the rate of progression of canopy closure. Although the interaction of cultivar by imazethapyr was nonsignificant, regression coefficients were less for Florunner, Southern Runner, Sunrunner, NC-7, and NC-V11 when treated with imazethapyr compared to each respective nontreated cultivar. Results from 1993 agree with 1992 canopy data that imazethapyr does slightly slow the progression of canopy closure. Previous research with imazapic showed a similar response, but reductions were larger (Richburg et al., 1994). All regression equations had correlation coefficients (r2) in the range of 0.81 to 0.96. Canopy width reductions by imazethapyr may delay canopy closure and lead to increased late season weed problems (Wilcut et al., 1994).
Peanut Grade and Yield-tifton, Ga
There was not a significant imazethapyr main effect or cultivar by imazethapyr interaction on percentage fancy pods, ELK or jumbo, medium, No. 1, sound splits, damaged, or other kernels, or TSMK (Table 2). There was a significant cultivar main effect on all above mentioned peanut grade components. Inspection of the data revealed that NC 7 had the highest fancy pod percentage followed by AgraTech VC 1 and NC-V11. Similar results were reported with imazapic for these varieties (Richburg et al., 1994). In general, virginia-type peanut had smaller percentage medium, No. 1, and other kernels than runner-types.
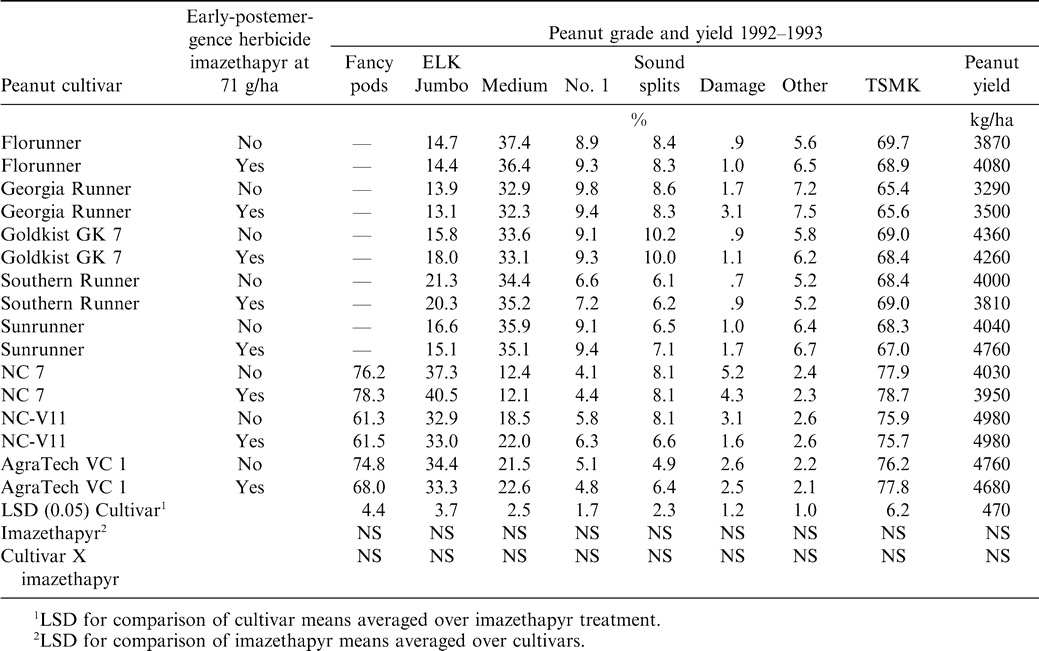
There was not a significant cultivar by imazethapyr interaction on percentage ELK or jumbo peanut kernels (Table 2). Cultivars NC 7, NC-V11, and AgraTech VC 1 had larger percentage ELK than Florunner, Georgia Runner, Goldkist GK 7, Southern Runner, and Sunrunner jumbo percentages. Virginia-type peanut have larger seed than runner-type peanut, consequently larger percentages of ELK compared to runner jumbo kernels. In contrast, previous research has shown that paraquat reduced ELK of NC 7 and Florigiant, both virginia-type peanut (Knauft et al., Florigiant 1990). Unlike previous research with imazapic, imazethapyr in this research did not reduce jumbo and ELK percentage of Florunner, Southern Runner, Sunrunner, and NC-V11 (Richburg et al., 1994). The only significant effect on TSMK was cultivar (Table 2).
The main effect of imazethapyr and the interaction of cultivar by imazethapyr were not significant on peanut yield (Table 2). Yield was influenced only by cultivar. The cultivar Georgia Runner yielded lower than other cultivars, otherwise no trends were evident. Peanut grade reductions and delays in maturity reported with imazapic (Richburg et al., 1994) and paraquat (Knauft et al., 1994) were not observed with imazethapyr.
Peanut Canopy Height and Width-yoakum, Tx
There was a significant cultivar and cultivar by imazethapyr effect on canopy height at 3 WAT (Table 3). Differences between peanut cultivars were expected since spanish- and runner-types were evaluated. The spanish cultivar Spanco had a higher canopy height when treated with imazethapyr at 3 and 6 WAT. Although not significant, reductions in canopy height at 3 WAT were found with Florunner, Pronto, Starr, and Tamspan 90, and with Pronto and Starr at 6 WAT when treated with imazethapyr.
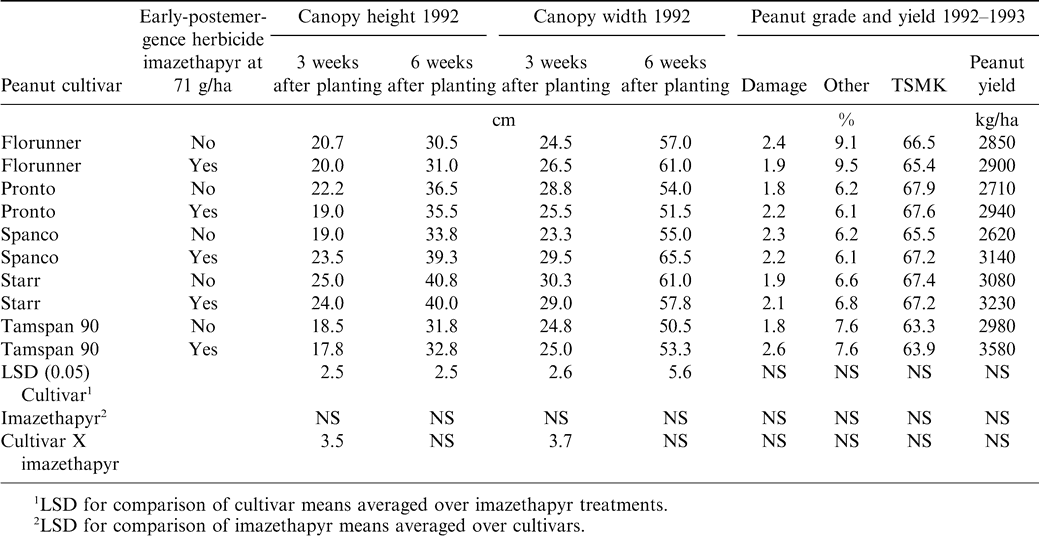
There was a significant cultivar effect at 3 and 6 WAT and cultivar by imazethapyr effect 3 WAT on canopy width (Table 3). As seen with canopy height, the cultivar Spanco had a larger canopy width when treated with imazethapyr.
Peanut Grade and Yield-yoakum, Tx
There were no significant effects on the percentage of damaged or other kernels, TSMK, and yield (Table 3). Although not significant, every cultivar evaluated had a higher yield when treated with imazethapyr. Previous research has shown that even though weeds were removed by hand, herbicide treated peanuts often have greater yields (Wehtje et al., 1991). This yield increase was attributed to less peanut disturbance and injury, which commonly occurs when hand weeding is required in plots with high weed densities.
Imazethapyr is the first herbicide registered in peanut to provide residual broadleaf and perennial nutsedge weed control (Richburg et al., 1995, 1996; Wilcut et al., 1991a, 1991b, 1994). Based on these results, a differential peanut cultivar response to imazethapyr should not be a problem. However, slightly slower canopy width closure in the Southeast may result in increased weed problems for Florunner, Southern Runner, Sunrunner, and NC-V11 peanut treated with imazethapyr. Conversely, the increase in canopy width of Florunner and Spanco peanut treated with imazethapyr may reduce late-season weed problems in Texas.
Acknowledgements
The authors express appreciation to Walter Harvey and Mary Amber Tiveron for their technical assistance in this research. This work was supported in part by State and Hatch funds (GEO00132) for the Georgia Agricultural Experiment Station, the Georgia growers through grants from the Georgia Agricultural Commodity Commission for Peanuts, State and Hatch funds for the Texas Agricultural Experiment Station, and the Texas Peanut Producers Board.
Literature Cited
Bailey W. A. , Wilcut J. W. , Spears J. F. , Isleib T. G. , and Langston V. B. 2000 Diclosulam does not influence yields in eight virginia market-type peanut (Arachis hypogaea) cultivars. Weed Technol 14 : 402 – 405 .
Bennett M. A. and Gorski S. F. 1989 Response of sweet corn (Zea mays) endosperm mutants to chloroacetamide and thiocarbamate herbicides. Weed Technol 3 : 475 – 478 .
Brecke B. J. 1989 Response of peanut cultivars to selected herbicide treatments. Proc. South. Weed Sci. Soc 42 : 28 .
Chiteka Z. A. , Gorbert D. W. , Stokes F. M. , Kucharek T. A. , and Knauft D. A. 1988 Components of resistance to late leaf spot in peanut. I. Levels of variability-implications for selection. Peanut Sci 15 : 25 – 30 .
Hammons R. O. 1976 Peanuts: Genetic vulnerability and breeding strategy. Crop Sci 16 : 527 – 530 .
Hardcastle W. S. 1974 Differences in the tolerance of metribuzin by varieties in soybeans. Weed Res 14 : 181 – 184 .
Hauser E. W. , Buchanan G. A. , Ethredge W. J. , Jellum M. D. , and Cecil S. R. 1976 interactions among peanut cultivars, herbicide sequences, and a systemic insecticide. Peanut Sci 3 : 56 – 62 .
Hauser E. W. , Buchanan G. A. , Harvey J. E. , Currey W. L. , Gorbet D. W. , and Minton N. A. 1981 Pesticide interactions with peanut cultivars. Peanut Sci 8 : 142 – 144 .
Holbrook C. C. and Kvien C. K. 1989 1988 cultivar census. Peanut Res 27 / 1 : 4 .
Johnson W. C. 1987 The hull scrape method to assess peanut maturity. Georgia Coop. Ext. Serv. Bull 958 .
Johnson W. C. , Holbrook C. C. , Mullinix B. G. , and Cardinia J. 1992 Response of eight genetically diverse peanut genotypes to chlorimuron. Peanut Sci 19 : 111 – 115 .
Johnson W. C. , Colvin D. L. , and Mullinix B. G. 1993 Comparative response of three peanut cultivars to multiple herbicide applications. Peanut Sci 20 : 17 – 20 .
Knauft D. A. , Colvin D. L. , and Gorbet D. W. 1990 Effect of paraquat on yield and market grade of peanut (Arachis hypogaea) genotypes. Weed Technol 4 : 866 – 870 .
Richburg J. S. , Wilcut J. W. , Culbreath A. K. , and Kvien C. K. 1994 Response of eight peanut (Arachis hypogaea) cultivars to AC 263,222. Peanut Sci 22 : 76 – 80 .
Richburg J. S. , Wilcut J. W. , and Eastin E. F. 1995 Weed management in peanut (Arachis hypogaea) with imazethapyr and metolachlor. Weed Technol 9 : 807 – 812 .
RichburgJ S. , Wilcut J. W. , and Vencill W. K. 1996 Imazethapyr systems for peanut (Arachis hypogaea L.). Peanut Sci 23 : 9 – 14 .
Rodriguez-Kabana R. , Backman P. A. , and Williams J. C. 1975 Determination of yield losses to (Sclerotium rolfsii) in peanut fields. Plant Dis. Rep 59 : 855 – 858 .
Smith A. E. and Wilkinson R. E. 1974 Differential absorption, translocation, and metabolism of metribuzin by soybean cultivars. Plant Physiol 32 : 253 – 257 .
Wehtje G. , Wilcut J. W. , McGuire J. A. , and Hicks T. V. 1991 Foliar penetration and phytotoxicity of paraquat as influenced by peanut cultivar. Peanut Sci 18 : 67 – 71 .
Wilcut J. W. , Walls F. R. , and Horton D. N. 1991a Imazethapyr for broadleaf weed control in peanuts (Arachis hypogaea). Peanut Sci 18 : 26 – 30 .
Wilcut J. W. , Walls F. R. , and Horton D. N. 1991b Weed control, yield, and net returns using imazethapyr in peanuts (Arachis hypogaea). Weed Sci 39 : 238 – 242 .
Wilcut J. W. , York A. C. , and Wehtje G. R. 1994 The control and interaction of weeds in peanuts. Rev. Weed Sci 6 : 177 – 205 .
Wixson M. B. and Shaw D. R. 1991 Differential response of soybean (Glycine max) cultivars to AC 263,222. Weed Technol 5 : 430 – 433 .
Notes
- 1Former graduate research assistant and 2Former Associate Professor Dep. of Crop and Soil Sciences University of Georgia Box 748 Tifton GA 31794-0748, and 3Research ScientistTexas Agric. Exp. Stn., Yoakum, TX 77995. Current address of J. W. Wilcut, Crop Science Department, Box 7620, North Carolina State University, Raleigh, NC 27695-7620. [^] *Corresponding author: john_wilcut@ncsu.edu.