

Introduction
Pod rot caused by Pythium spp. and Rhizoctonia spp. affects peanut yield and quality. In Oklahoma, 43% of 37 peanut fields sampled had pod rot, with the disease incidence in the range of 5 to 37% (Filonow et al., 1988). Pod rot was found to cause considerable losses in Israel (Frank, 1968). In North Carolina, pod rot was estimated at 6 to 7% in 1995 and 1996 (Hollowell, et al., 1998). In both North Carolina (Hollowell et al., 1998) and the High Plains of Texas (Wheeler et al., 2006), Rhizoctonia spp. and Pythium spp. were the most common pathogens isolated from peanuts with pod rot symptoms.
Peanut producers with pod rot problems caused by either Pythium spp. alone, or by a combination of Pythium spp. and Rhizoctonia, can spend > $250/ha/season in fungicide costs in an effort to minimize losses. Producers rely heavily on fungicides as the primary means of reducing pod rot caused by fungi and oomycetes and other fungal disease problems. Very few cultivars are recognized to have pod rot tolerance or resistance (Baughman and Dotray, 2007). Calcium has been found to reduce pod rot incidence in soils where calcium is deficient (Csinos and Gaines, 1986).
Irrigation can affect diseases caused by fungi and oomycetes. In Israel, pod rot developed in well-aerated sandy soils, which were irrigated frequently (Frank and Ashri, 1985). Under aerated conditions, both wet and dry conditions favor the growth of R. solani (Rotem and Palti, 1969). Infection and disease development of soil rot of tomato caused by R. solani was favored when soil moisture was at 60% of the water holding capacity of soil (mixture of sand and peat) when compared to 20, 40, and 100% levels (Gonzalez and Owen, 1963). Damage caused by Pythium spp. on peanut pods increased with frequent short irrigations compared with heavier irrigations for longer time periods (Frank, 1967). Incidence of pod rot induced by Sclerotinia minor and P. myriotylum was higher in sprinkler irrigated plots than in plots that were not irrigated (Porter et al., 1987).
In the semiarid Southern High Plains of Texas, rainfall during the pod forming period may be limited, so most producers use overhead center pivot irrigation to provide water for good crop growth and suitably moist conditions for pegging and pod development. Depending on weather conditions, peanuts grown in West Texas require about 50 to 70 cm of water during the growing season for optimum peanut yields. Application of 2.5 to 4 cm of water weekly, split into two applications is recommended during peak blooming period (from early July to late August) (Baughman and Dotray, 2007). However, deficit irrigation due to limited irrigation capacities is common in this region, as pumping capacities of irrigation wells are often insufficient to meet peak water demand for peanuts. Hence, higher irrigation rates are generally associated with higher yields in peanuts for this region. However, there is a poor understanding of the tradeoff between higher irrigation rates and pod rot. The objectives of this study were to investigate the impact of irrigation rates and fungicides on kernel damage, yield and net returns on peanut production in the Southern High Plains of Texas, in a field with a history of Pythium and Rhizoctonia pod rot.
Materials and Methods
Field studies were conducted in Gaines County, TX (Longitude = −102.745, Latitude = 32.903, altitude = 1039 m). The soil type was Brownfield loamy sand (loamy, mixed, superactive, thermic Arenic Aridic Paleustalfs) of 81% sand, 4% silt, and 15% clay, OM 0.1%, and pH 7.8. Tests were conducted in the two innermost spans of the center pivot irrigation system with rows planted in circles. This area was kept in continuous peanuts from 2000 to 2004. Ethalfluralin (Sonalan; 1.75 L/ha) was used as a preplant herbicide. Post-emergence herbicides (including Benzothiazol/Diphenylether, Storm; Bentazon, Basagran) were used on peanuts as needed. No insecticides were applied to peanuts. The center pivot system nozzle package was modified to apply three irrigation rates consisting of a base (B) rate, B + 33% rate, and B − 33% rate. The base rate was designed to meet approximately 75% of the crop needs based on estimated crop evapotranspiration (ETc) (Jensen et al., 1990), and subject to the limits of pumping capacity for that center pivot system. In most years, the pumping capacity was insufficient during the months of July and August when crop water demand is highest. In 2002, the irrigation amounts applied were 47, 71, and 95 cm for low, medium, and high rates of irrigation, and in-season rainfall totaled 9 cm. In 2003, the irrigation amounts applied were 36, 53, and 71 cm for low, medium, and high rates of irrigation, and in-season rainfall totaled 24 cm. In 2004, the irrigation amounts applied were 37 and 46 cm for medium and high rates of irrigation, and in-season rainfall totaled 44 cm. Daily reference evapotranspiration (ETo) and peanut evapotranspiration (ETc) were calculated from a weather station located near Lamesa, TX (http://txhighplainset.tamu.edu). This weather station website is no longer available as of September 2010.
In all 3 yrs of the study, cv. FlavorRunner 458 was planted on 91 cm row spacing. Bradyrhizobium inoculant was applied. Planting dates were 25 April 2002, 30 April 2003, and 26 April 2004.
During June each year, the center pivot irrigation system nozzle package (Senninger LDN low drift spray nozzles, Senninger Irrigation, Clermont, FL, www.senninger.com) was modified to apply the three irrigation rates. Spray nozzles were operated at 41 kPa pressure by using pressure regulators. Nitrogen fertilizer at 67.2 kg N/ha/season was supplied by three applications of 22.4 kg N/ha during the growing season. The first application went out preplant using 30-24-0-0; the second application went out in late June using 28-0-0-5; and the third application went out in late July using 28-0-0-5.
Irrigation rate treatments were arranged in a randomized complete block design with three replications. Irrigation treatments were 14 rows wide in the first span (replication 1), and seven rows wide in the second span (replications 2 and 3) under the center pivot irrigation system. Each irrigation treatment was applied to the same circular rows of the entire circle. Fungicide plots were two-rows wide by 4.6 m long and were replicated in each irrigation plot [i.e. nine replications for each fungicide treatment (three irrigation rates × three replications) within the test area]. Fungicide treatments consisted of a non fungicide check, azoxystrobin (Abound FL) at 0.45 kg ai/ha applied at 60 and 90 days after planting, and metalaxyl (Ridomil Gold EC) applied at 0.24 kg ai/ha applied at 90 days after planting in 2002, and applied at 60 and 90 days in 2003 and 2004. Fungicides were applied with a CO2-pressurized back pack sprayer at 110 kPa. The fungicide applications were applied using a hand-held boom with teejet® nozzles (8002VS) spaced at one nozzle per row and the rate of 85 L water/ha. In all years, water (2805 L/ha) was applied through a sprinkler device (developed by T. Wheeler and D. Porter, Fig. 1) to the plots within 1 hr after the fungicide was applied to mimic chemigation.

Device used to apply water over the top of fungicide treated plots, to simulate chemigation. The device was constructed by T. Wheeler and D. Porter.
To determine the causal agents of diseased pods in 2003 and 2004 (but not in 2002), five plants were removed from each plot at 60 days after planting (before the first fungicide application). Pods with rot or discoloration symptoms (four pods/plot were used if sufficient diseased pods were available) were washed under running tap water for 2 min and then dried in a flow of sterile air under a laminar flow hood for 3–4 hr. Lesions were then placed on water agar (1.5%) and a Pythium selective media (Mircetich, 1971). Fungal and oomycete growth from these plates was transferred to potato dextrose agar and identified to genus.
Peanuts were inverted using a two-row digger on 18 October 2002, 6 October 2003, and 11 October 2004. After plants had dried for ≥1 wk, pods were harvested using a small plot thresher and weighed. A 250-g sample from each plot was graded. The sample was shelled and all damaged kernels were weighed to obtain percent damaged kernels (DK). Shelled peanuts were sieved (0.64 × 1.9 cm openings) to obtain sound mature kernels (SMK), sound splits (SS), foreign material, and other kernels (OK = other kernels that were small or defective, minus SS). Grade was the combined percentage of SMK and SS.
Runner peanut values were calculated by ($5.34 × percent Grade) + ($1.54 × percent OK)/1000 kg peanuts (prices were based on the 2009 peanut loan schedule, obtained from Golden Peanut Company, Brownfield, TX), minus deductions and fungicide application costs. Deductions for damaged kernels cost $3.74/1000 kg peanuts for samples with 1.5 to 2.49% damage, and for higher amounts of damage, the peanuts were valued at 35% of their calculated value for grade and other kernels (based on a peanut loan schedule for the 2009 crop, obtained from Golden Peanut Company). Deductions were also acquired for %SS that were > 4.49%. For each additional percentage of SS, the value was reduced by $0.88/1000 kg peanuts based on a peanut loan schedule for the 2009 crop. Azoxystrobin cost $146.75/ha for two broadcast applications, and metalaxyl cost $120.47/ha for two broadcast applications. Peanut pod yield/ha was calculated based on the plot weights, and an overall value (grade × price - deductions)/ha was calculated. Net return ($/ha) was calculated from the overall value (yield × price), minus pumping and fungicide costs. Pumping costs for the irrigation treatments were calculated based on a depth to the groundwater table of 39.4 m, typical for the vicinity of the field studied. The operating pressure for the pivot was approximately 138 kpa and the pivot was nozzled at 2120 L/min. The electricity cost was estimated at $0.073/kWh, and the pumping plant efficiency was assumed to be 46.2%, an average based upon surveys of pumping plants in the area (Fipps and Neal, 1995). The pumping cost given these assumptions would be $0.0261/m3 ($2.68/acre-inch).
Isolations were made as described previously using pods with lesions collected immediately after inverting plots. Pods were stored in paper sacks until isolations were conducted. Eight pods/plot were used in the isolation procedures at harvest if sufficient number of diseased pods were available.
Data were analyzed using PROC MIXED (SAS version 9.1, Cary, NC) on DK, grade, yield (kg/ha), and the net return/ha. The analysis was based on a split plot design with irrigation rate (IrrRate) as the main plot and fungicide treatment (TRT) as the subplot. The model statement included IrrRate, TRT, and their interaction. The initial analysis was conducted on all three data sets combined. The random statement had year, year nested within replication [REP(year)], year × IrrRate, REP × IrrRate (year), year × TRT, and year × IrrRate × TRT. However, the results were inconsistent among years, particularly in regards to the random factor REP × IrrRate(year) which had a significant effect, so each year was analyzed separately. In these analyses, the model statement was the same as for the overall analysis, but the random statement contained REP, and REP × IrrRate. In 2004, there were no B − 33% irrigation rate that was treated with fungicides so only the B and B + 33% irrigation rates were included in that analysis. Factors were considered significant if P ≤ 0.05.
In order to look at the relative importance of water and fungicide application to yield, regression analysis was performed with these factors. Fungicide application was coded as 1 if azoxystrobin was used and 0 if metalaxyl or no-fungicide was used. The linear and quadratic function of water (cm of irrigation plus rainfall during the growing season) was also used as a factor. Initially the linear, quadratic, and interaction terms were examined across all three years, using PROC RSREG (SAS) to identify the significant terms. Then regression analysis was used to develop the final equation with those terms that were significant with PROC RSREG. Factors were included that were significant at P ≤ 0.05.
The isolation frequency of pathogens (Rhizoctonia spp. and Pythium spp.) at 60 days after planting, as affected by IrrRate and presence of Pythium spp. or Rhizoctonia spp. was determined using PROC MIXED (SAS). The data for the analysis consisted of each pod separately, and positive for either Pythium spp., Rhizoctonia spp., or neither. In none of the samples were both organisms isolated from the same lesion. The model statement included IrrRate and fungal type (i.e. the incidence of Pythium spp. isolation and of Rhizoctonia spp. isolation), and their interaction with the chi-square option used to determine significance at P = 0.05. The Satterthwaite method was used to calculate degrees of freedom and the PDIFF option was used to determine differences between treatments at P = 0.05. At harvest, a similar analysis was conducted, except fungicide treatment (TRT) was also included in the model statement as a main effect, and with 2-way and 3-way interactions. The random states for both analyses were similar to that done for yield, and initially included year as a factor. However, as was found with the previous analyses, the REP × IrrRate (year) was significant, so each year was analyzed separately.
Results and Discussion
Mean air temperature in 2002 was ca. 26 °C during June, July, and August, and then became unseasonably warm in September with mean ca. 28 °C (Fig. 2A). Warm temperatures were coupled with low levels of rainfall (Fig. 2B) during all months of the growing season. In 2003, air temperature was highest during July and August, as is typical in this region and very cool during September (Fig. 2A). Rainfall was low during July through August 2003, when plant water demands were highest. In 2004, air temperature was unusually cool during August and rainfall was unusually high during September (Fig. 2A,B).
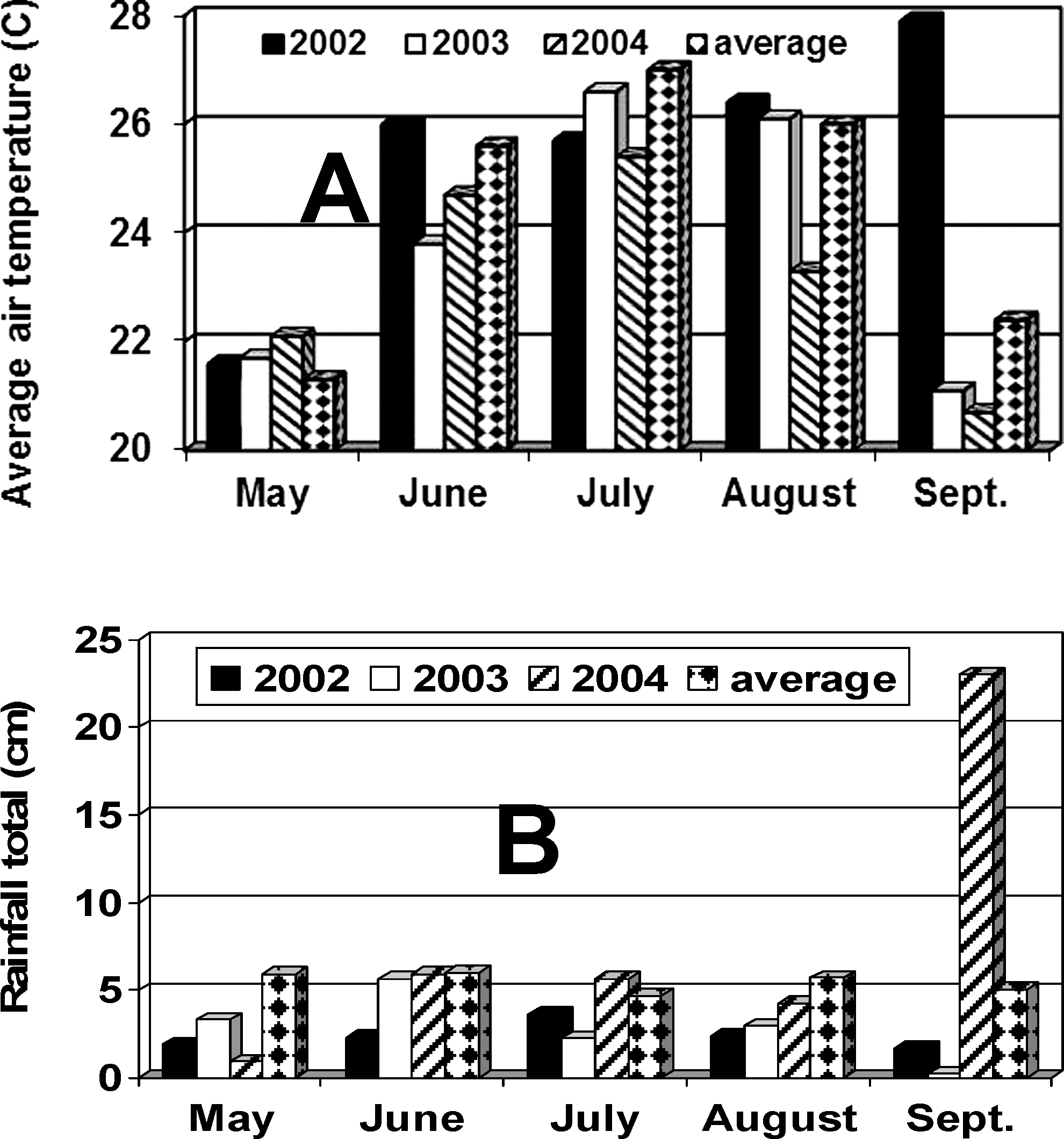
Average air temperature (A) and rainfall (B) for 2002–2004 collected with a weather station at the test site. The long term average for air temperature and rainfall are also included in the Figure
 (obtained from the National Climate Data Center, NOAA).
(obtained from the National Climate Data Center, NOAA).
In 2002 and 2004, there was no effect of irrigation on DK. In 2003 there was an interaction between irrigation rate and fungicide treatment, with respect to DK. At the B − 33% irrigation rate there were more (P < 0.01) DK associated with the metalaxyl (1.4% DK) and no-fungicide (1.5% DK) treated plots than the azoxystrobin (0.1% DK) treated plots. At the B irrigation rate, there were no effect of fungicide treatments on DK (average of 0.3% DK), and at the B + 33% irrigation rate, there were more (P < 0.01) DK associated with the metalaxyl (2.0% DK) treated plots than the azoxystrobin (0.3% DK) or no-fungicide (0.7% DK) treated plots.
Irrigation rate did not significantly (P ≤ 0.05) impact peanut grade except in 2004, where kernels from plots with the B irrigation rate had a higher grade (79) than did kernels from plots in the B + 33% irrigation rate (76). Irrigation rate did affect (P < 0.04) yield in 2 of 3 yr (Table 1). Plots from the B + 33% irrigation rate had higher yields in 2002 and 2003 than plots from the B and B − 33% irrigation rates (Table 1). Plots from B − 33% irrigation rate had lower yields than plots from the B irrigation rate in 2002 (Table 1). Irrigation rate did not affect the net return of the crop/ha in any year (P > 0.37 in all years). Net return factored in pumping costs for the irrigation as well as value of the peanuts and fungicide costs.
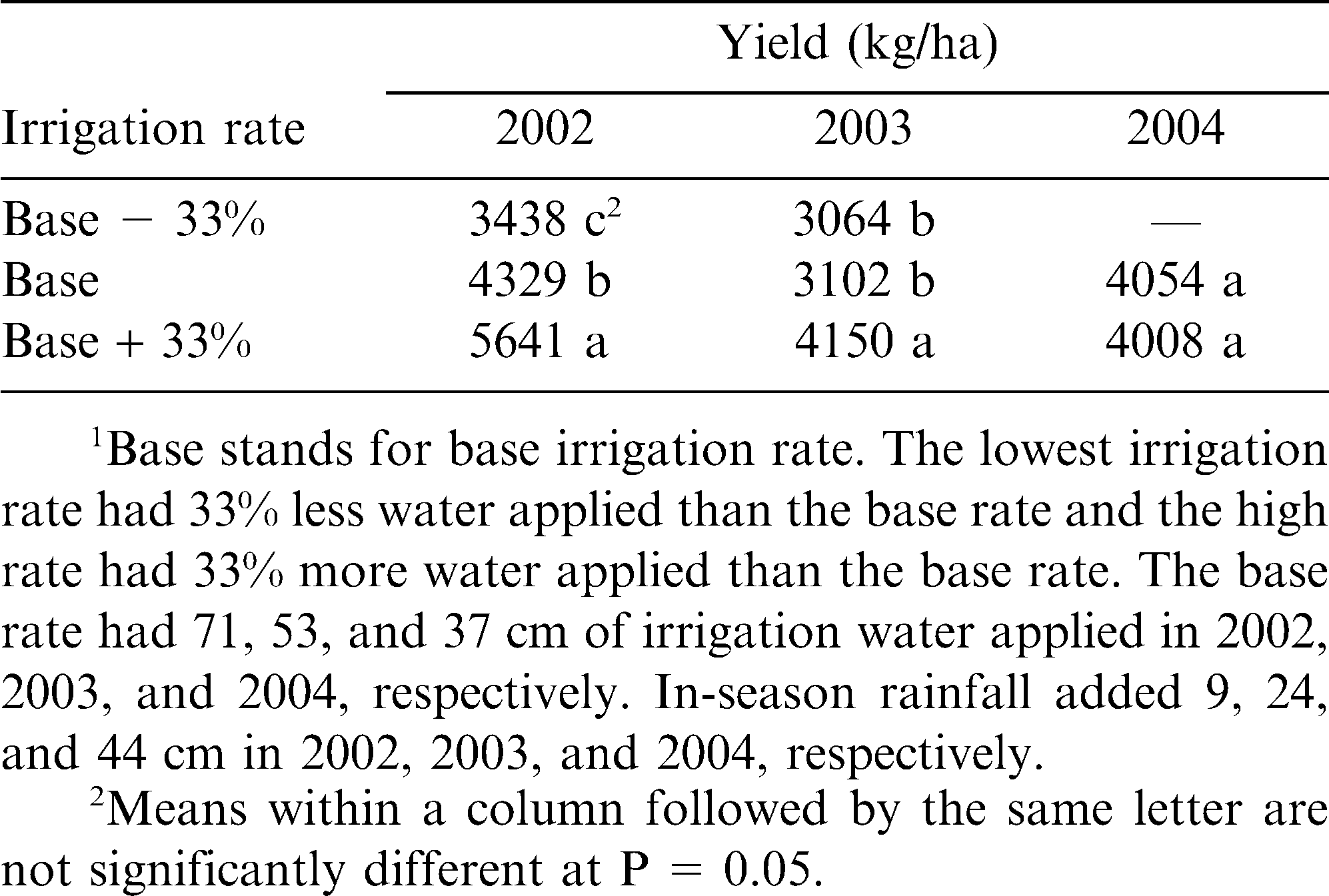
Effect of irrigation rate1 on yield in 2002, 2003, and 2004 at a field in Gaines County, TX.
Plots treated with the fungicide azoxystrobin had fewer DK than plots with no-fungicide or plots treated with metalaxyl in all 3 years (Table 2). Metalaxyl treated plots had similar DK (2002, 2004) or more DK (2003) than the no-fungicide plots in all years. Peanut grades were higher for plots treated with azoxystrobin than for no-fungicide plots or plots treated with metalaxyl in all 3 years (Table 2). Yields were higher in the plots treated with azoxystrobin than no-fungicide plots in all 3 yr. Metalaxyl treated plots had higher yield than the no-fungicide treated plots in 1 of 3 yr (2003). Net return/ha was significantly (P < 0.05) higher for plots treated with azoxystrobin than plots receiving no-fungicide or metalaxyl treated plots in 2 of 3 yr (Table 2).
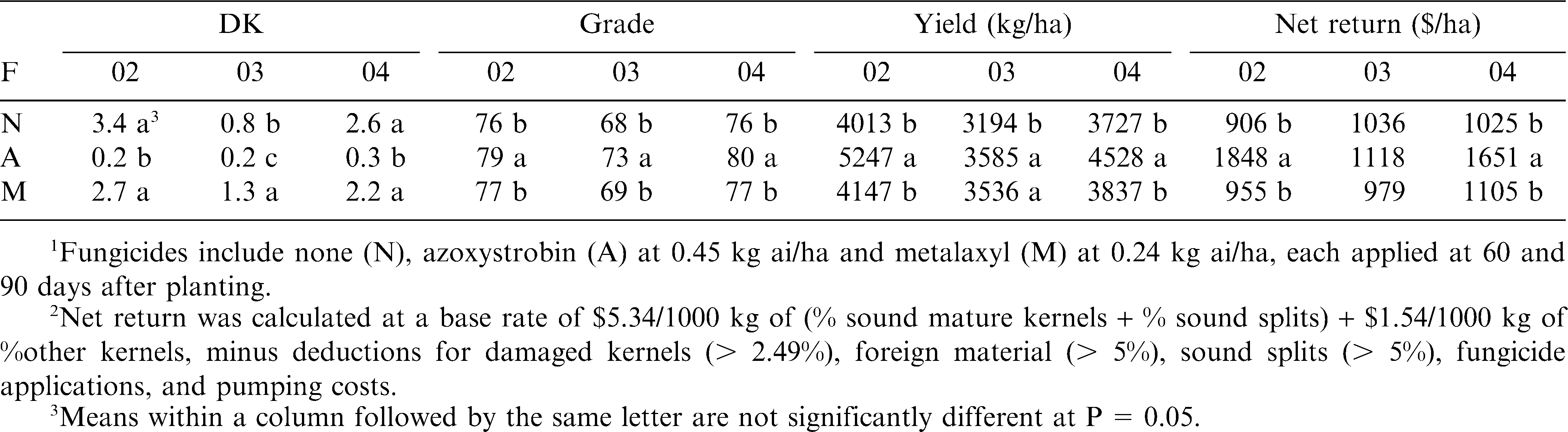
Effect of fungicides (F1) on percent damaged kernels (DK), yield, and net return2 of the peanut crop in 2002 (02), 2003 (03), and 2004 (04) at a field in Gaines County, TX.
Yield was predicted with a quadratic function for water (irrigation plus rainfall in cm during the season) and linear function for fungicide (1 = azoxystrobin, 0 = metalaxyl or no-fungicide treatment) (P < 0.01). The interaction term between water and fungicide was not significant (P = 0.88). The model fitted by regression analysis was:

Azoxystrobin application resulted in an average yield increase of 761 kg/ha based on this model. The average value of peanuts in azoxystrobin plots was $416.85/1000 kg peanuts. The cost of the broadcast treatment was $146.75/ha. The net return of the treatment after subtracting fungicide costs was $170/ha. The net return of irrigation to the crop is more complicated to calculate due to the quadratic function and because some of the water was rainfall. To compare the relative importance between fungicide and irrigation, azoxystrobin added 761 kg/ha at a cost of $146.74/ha. To get a similar increase in yield, it would be necessary to go from 56 cm water (the lowest amount in the study) to 86 cm water (Fig. 3). The cost of pumping 30 cm of water/ha is $76.75/ha, given the stated assumed conditions for that site (depth to groundwater, irrigation system pressure, given efficiency, etc.). Other locations would note different costs of pumping, and these values can vary substantially. So, the cost of increasing yield due to water at this site was 52% of the cost of increasing yield a similar increment due to fungicide application. However, with fungicide application, there is a limit to the response, while for water there was no limit to the increase that occurred over the range of the study. However, as the water amount approaches the maximum that the crop required, there would be an expectation of diminished returns. The maximum water applied during the growing season was > 100 cm, which greatly exceeds the published maximum peanut ETc (Bandyopadhyag et al., 2005; Kheira, 2009). The fact that yield was still increasing in response to water in our study at this amount suggests that there may be greater losses to evapotranspiration and deep percolation than found in published studies, and peanut evapotranspiration models may not be a good fit for this region.
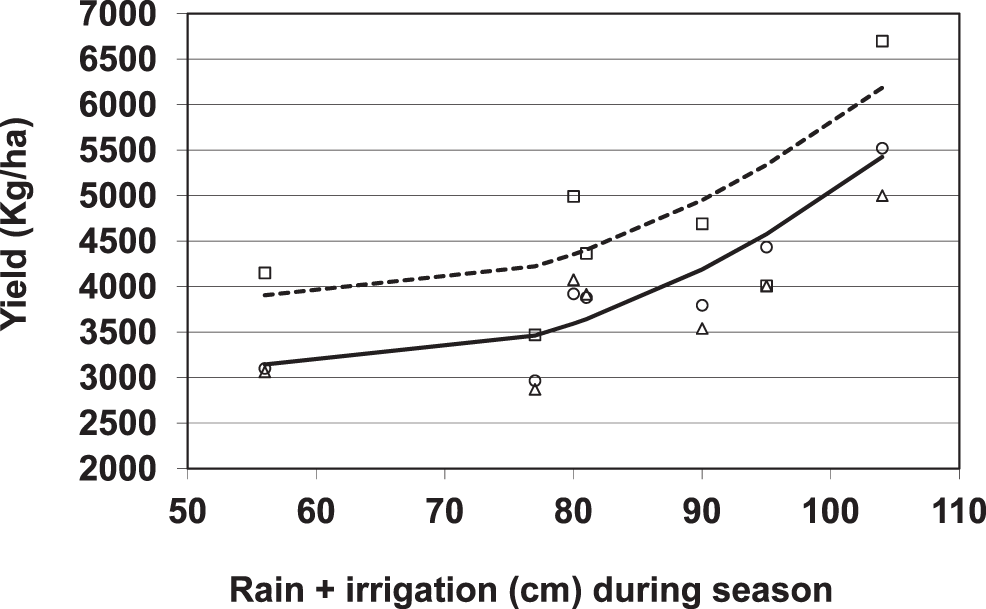
Effect of water (rainfall plus irrigation) and azoxystrobin on peanut yield from 2002–2004. Water was in cm, and if azoxystrobin was applied then X = 1, otherwise, X = 0. Model predicted was: Yield (kg/ha) = 7473.3 + 760.9(X) − 144.5(Water) + 1.2(Water2), R2 = 0.50, P < 0.01 for all estimated parameter in the model. Average yield for a given water level for plots treated with axoxystrobin (□), metalaxyl (○), and no-fungicide (△). The line predicting yield in the absence of azoxystrobin (—) and the line predicting yield in the presence of azoxystrobin (– – – –).
Rhizoctonia spp. and Pythium spp. were isolated from diseased pods, but more Rhizoctonia spp. was found than Pythium spp., both at midseason and harvest in 2003 and 2004 (Tables 3 and 4). Irrigation rate did not have a consistent effect on isolation frequency of either Rhizoctonia spp. or Pythium spp. (Tables 3 and 4). However, when azoxystrobin was applied, then isolation frequency of Rhizoctonia spp. was low. The isolation frequency and the performance of azoxystrobin compared with metalaxyl and the no-fungicide treatments suggest that the primary pod rot pathogen in the field was caused by Rhizoctonia. Metalaxyl, which only protects against the oomycetes (Hoy and Schneider, 1988; Stone et al., 1987), did not perform as well as azoxystrobin, which protects against many fungi and oomycetes, including some activity against Pythium spp (Besler et al., 2003; Grichar et al., 2000). In this field, Pythium spp. was probably not as important a component in the pod rot complex as was Rhizoctonia.

Effect of irrigation rate on frequency of isolation of Pythium spp. and Rhizoctonia spp. from discolored and rotted pods at 60 days after planting.
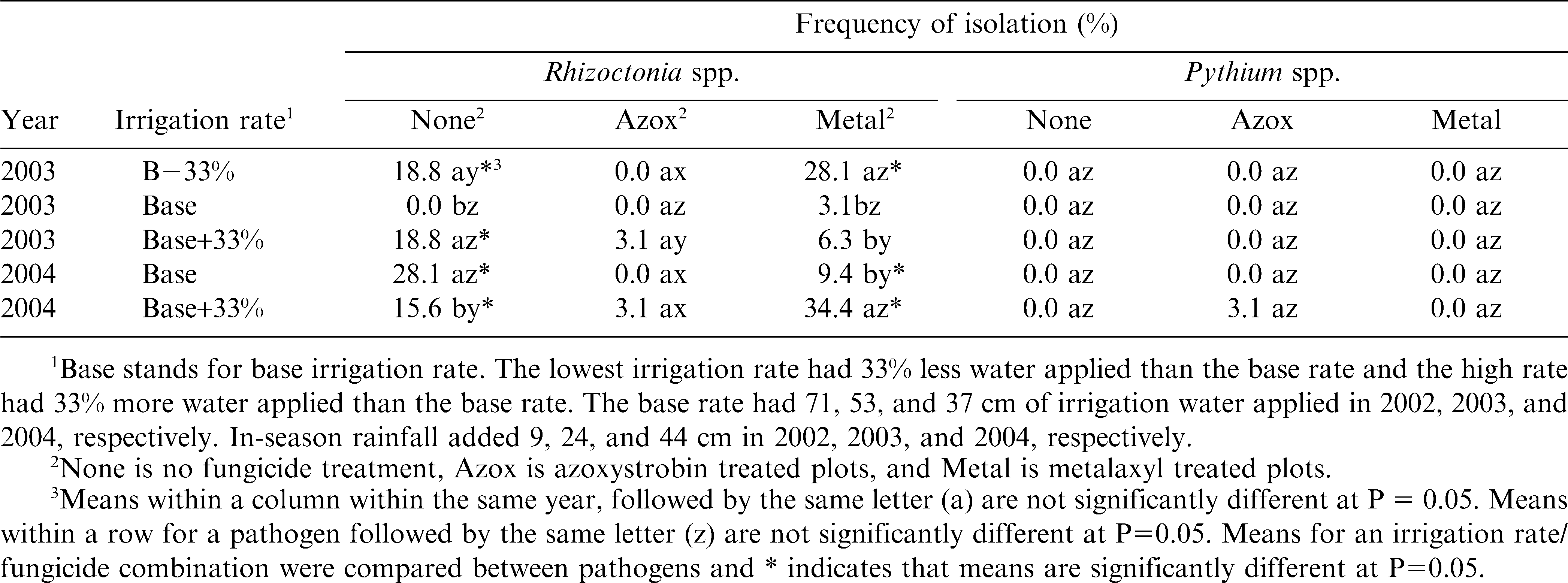
Effect of irrigation rate and fungicide treatment on frequency of isolation of Pythium spp. and Rhizoctonia spp. from discolored and rotted pods at harvest during 2003 and 2004.
Producers in this region associate higher irrigation capacity with higher yield. This relationship was also consistent with our results. However, the increase in yield did not result in higher net return. Soilborne diseases caused by fungi and oomycetes are often more severe under wet conditions (Cook and Papendick, 1972; Duniway, 1979). Damaged kernels, which can be caused by Pythium spp. and Rhizoctonia spp. were generally not associated with irrigation rate in these studies. Yield, however, was affected by irrigation rate in 2 of 3 yr and in 2004 where irrigation rate was not significant, the irrigation treatment totals only differed by 9-cm applied over the entire season.
DK was strongly affected by fungicide treatment. Azoxystrobin greatly reduced the amount of DK and improved yields in all 3 yr of the test. Metalaxyl was ineffective on DK in all 3 yr.
Losses due to pod rotting organisms can reduce harvestable yield by destroying pegs, and pods may detach before they can be harvested. Therefore, DK does not include all peg and pod disease. Yield and net return (including penalties for DK) may be a better measure, since it would represent the loss in pods from peg rot before pod formation or peg rot that detached pots. Since azoxystrobin did positively impact percent damaged kernels every year, we conclude that irrigation rate had minimal impact on disease compared with application of azoxystrobin. Increasing amounts of irrigation led to higher yield in 2 of 3 yr without an increase in DK. If a producer from this region, or a region with a similarly high evapotranspiration rate coupled with low rainfall, chooses to maximize yield with irrigation, there is no evidence that it will lead to more kernel damage caused primarily by Rhizoctonia spp. However, use of a fungicide like azoxystrobin to protect pods is recommended, particularly if there is any history of pod rot.
Acknowledgements
The work presented was supported by the Western Peanut Growers Association, Texas AgriLife Research, Texas AgriLife Extension Service, and a grant from the Texas AgriLife Research Precision Agriculture Program (now known as the Texas AgriLife Research Cropping Systems Program) at Texas A&M University.
Literature Cited
Baughman T and Dotray P (eds.) 2007 Texas Peanut Production Guide Texas Cooperative Extension , College Station, TX http://peanut.tamu.edu/pdfs/productionguide07.pdf. Accessed September 1, 2009 .
Bandyopadhyag P.K Mallick S and Rana S.K 2005 Water balance and crop coefficients of summer-grown peanut (Arachis hypogaea L.) in a humid tropical region of India Irrig. Sci. 23 : 161 – 169 .
Besler B.A Grichar W.J Brewer K.D and Baring M.R 2003 Assessment of six peanut cultivars for control of Rhizoctonia pod rot when sprayed with azoxystrobin or tebuconazole Peanut Sci. 30 : 49 – 52 .
Cook R.J and Papendick R.I 1972 Influence of water potential of soils and plants on root diseases Annu. Rev. Phytopathol. 10 : 349 – 374 .
Csinos A.S and Gaines T.P 1986 Peanut pod rot complex: A geocarposphere nutrient imbalance Plant Dis. 70 : 525 – 529 .
Duniway J.M 1979 Water relations of water molds Annu. Rev. Phytopathol. 17 : 431 – 460 .
Filonow A.B Melouk H.A Martin M and Sherwood J 1988 Effect of calcium sulfate on pod rot of peanut Plant Dis. 72 : 589 – 593 .
Fipps G and Neal B 1995 Texas Irrigation Pumping Plant Efficiency Testing Program. Final Report to the State Energy Conservation Office Dept. of Biological and Agricultural Engineering, Texas AgriLife Extension Service , College Station, TX. Accessed at: http://itc.tamu.edu/documents/Pumping Plant Eff.pdf.
Frank Z.R 1967 Effect of irrigation procedure on Pythium rot of groundnut pods Plant Dis. Rep. 56 : 414 – 416 .
Frank Z.R 1968 Pythium pod rot of peanut Phytopathology 58 : 542 – 543 .
Frank Z.R and Ashri A 1985 Pythium pod rot reduction in peanuts, as affected by breeding for resistance and by disease level Phytoparasitica 13 : 41 – 45 .
Gonzalez L.C and Owen J.H 1963 Soil rot of tomato caused by Rhizoctonia solani Phytopathology 53 : 82 – 85 .
Grichar W.J Besler B.A and Jaks A.J 2000 Use of azoxystrobin for disease control in Texas peanut Peanut Sci. 27 : 83 – 87 .
Hollowell J.E Shew B.B Beute M.K and Abad Z.G 1998 Occurrence of pod rot pathogens in peanuts grown in North Carolina Plant Dis. 82 : 1345 – 1349 .
Hoy J.W and Schneider R.W 1988 Role of Pythium in sugarcane stubble decline: Effects on plant growth in field soil Phytopathology 78 : 1692 – 1696 .
Jensen M.E Burman R.D and Allen R.G 1990 Evapotranspiration and Irrigation Water Requirements, ASCE Manuals and Reports on Engineering Practices No. 70. ASCE New York .
Kheira A.A.A 2009 Macromanagement of deficit-irrigated peanut with sprinkler irrigation Agric. Water Man. 96 : 1409 – 1420 .
Mircetich S.M 1971 The role of Pythium in feeder roots of diseased symptomless peach trees and in orchard soils in peach tree decline Phytopathology 61 : 357 – 360 .
National Climatic Data Center, NOAA, Accessed at http://cdo.ncdc.noaa.gov/cgi-bin/climatenormals/climatenormals.pl .
Porter D.M Wright F.S and Powell N.L 1987 Effects of sprinkler irrigation on peanut diseases in Virginia Plant Dis. 71 : 512 – 515 .
Rotem J and Palti J 1969 Irrigation and plant diseases Annu. Rev. Phytopathol. 7 : 267 – 288.
Stone J.R Verma P.R Dueck J and Westcott N.D 1987 Bioactivity of the fungicide metalaxyl in rape plants after seed treatment and soil drench applications Canad. J. Plant Path. 9 : 260 – 264 .
Wheeler T.A Howell C.R Cotton J and Porter D 2006 Pythium species associated with pod rot on west Texas peanuts and in vitro sensitivity of isolates to mefenoxam and azoxystrobin Peanut Sci. 32 : 9 – 13 .
Notes
- Texas AgriLife Research, Lubbock, TX 79403
- Formerly at Division of Agriculture, West Texas A&M University, Canyon, TX 79016
- Texas AgriLife Extension Service, Lubbock, TX 79403
- Texas A&M University at Kingsville, TX 78363
- Formerly at the Texas AgriLife Research, Lubbock, TX 79403, now deceased *Corresponding author, email: ta-wheeler@tamu.edu
Author Affiliations