

Introduction
Sterile brachytic peanut ( Arachis hypogaea L.) plants have previously been reported (Ashri 1968, Coffelt and Hammons 1972, Hull 1937). These plants have been observed in F2 infraspecific cross populations between the two subspecies, ssp. hypogaea and ssp. fastigiata (Hammons 1973). The inheritance of sterile brachytic plants has been reported to be controlled by one recessive gene (Hull 1937), two duplicate recessive genes (Ashri 1968, Patel et al., 1936), and four duplicate recessive genes (Coffelt and Hammons 1972).
Brachytic plants are sterile with no flowers, short stem internodes, and clustering of the four leaflets without apparent rachis (Ashri 1968). However, similar sterile dwarf plants were found recently but had apparent rachis and petiole leaflet trait and presence of flowers. These traits appear to be the main distinguishing difference between the sterile brachytic and the sterile dwarf plants. The objective of this genetic study was to determine the inheritance of these two sterile dwarf genotypes found within segregating F2 populations, and their implications for peanut breeding and genetics.
Materials and Methods
Two cross combinations were made in the greenhouse between and within subspecies of Arachis hypogaea L. C1721 was an infraspecific cross between A. hypogaea ssp. hypogaea cv. ‘Georgia-11J' (Branch 2012) X A. hypogaea ssp. fastigiata cv. ‘Georgia Valencia' (Branch 2001). Whereas, C1813 was a cross made within A. hypogaea ssp. hypogaea and involved a three-way cross between [‘Georgia-06G' (Branch 2007) X GA 032913, a sister-line of ‘Georgia-09B' (Branch 2010)] X ‘Tifguard' (Holbrook et al., 2008).
Seed of F1, F2, and F3 populations were space-planted 30.5cm apart in field nursery plots at the agronomy research farm near the University of Georgia, Coastal Plain Experiment Station, Tifton Campus. Each year, plots consisted of two rows with variable length depending upon number of seed x 1.8m wide, and were planted on a Tifton loamy sand soil type (fine-loamy, siliceous, thermic, Plinthic Kandiudult). Recommended cultural practices with irrigation were followed throughout the growing season. Individual plants were harvested near optimum maturity based upon days after planting and above-ground appearance. After harvest, peanut pods were dried with forced warm air to approximately 6% moisture content before weighing and shelling.
Phenotypic classification was based on individual plants after digging and inverting. Segregation data was analyzed by chi-square analysis for goodness-of-fit (P≤0.05) to expected genetic ratios (Strickberger 1968).
The starch content of pollen grains was estimated by staining pollen deposited onto microscope slides with Fisher brand 2% iodine-potassium iodide for 10 min prior to observation. Pollen stainability was estimated by classifying pollen grains within the field of view at x100 magnification as viable (stained and regularly shaped) or inviable (not stained or irregularly shaped).
Results
Each F1 plant from both crosses (C1721 and C1813) was classified as normal. This indicates that both sterile brachytic and sterile dwarf were recessive to normal plant genotypes.
The F2 segregation from C1721 (between subspecies hypogaea and fastigiata) showed an acceptable fit for both a 243 normal to 13 sterile brachytic and a 15 normal to 1 sterile brachytic expected ratio, respectively (Table 1). Total, summed, and homogeneity chi-square values were also found acceptable for each of the two expected ratios. These results agree with the two duplicate recessive gene model of Ashri (1968) and Patel et al. (1936) as well as the four homozygously recessive genetic model proposed by Coffelt and Hammons (1972) for inheritance of sterile brachytic genotypes. However, these results disagree with the single recessive gene model proposed by Hull (1937) for this trait.
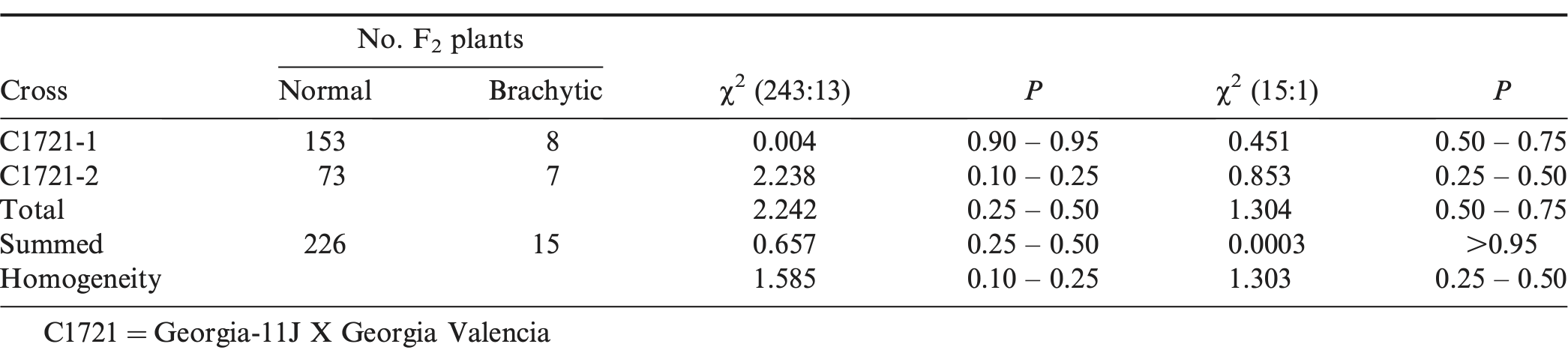
F2 segregation for number of normal and sterile brachytic plants from peanut cross combination between subspecies hypogaea and fastigiata.
The F2 segregation from C1813 (within subspecies hypogaea) showed a 3 normal to 1 sterile dwarf expected genetic ratio (Table 2). Segregation only occurred within one of four F1:2 families from this cross combination. The F3 results confirmed a monogenic model with an acceptable fit to a 2 segregating to 1 non-segregating F2:3 progeny rows (χ2 = 0.98, P = 0.25 – 0.50). The gene symbol, sdw, is proposed for this new sterile dwarf peanut genotype.
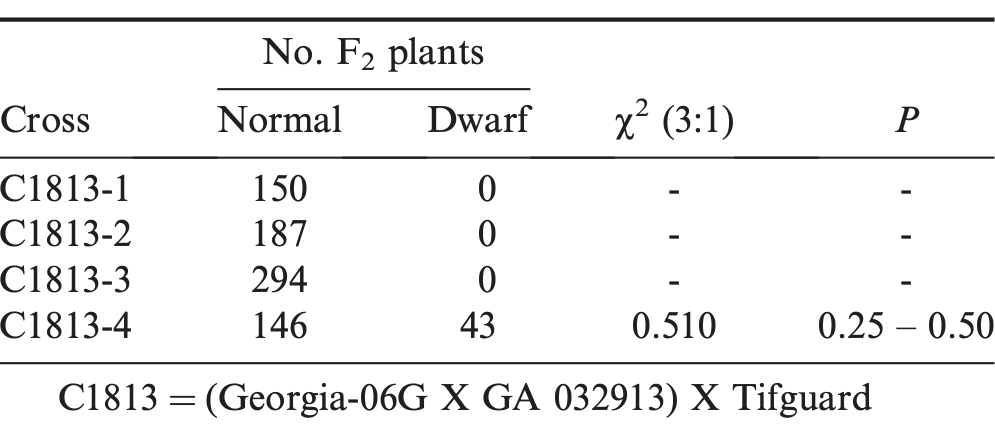
F2 segregation for number of normal and sterile dwarf plants from peanut cross combination within subspecies hypogaea
Discussion
These findings suggest that even though both the sterile dwarf and the sterile brachytic plants are somewhat similar phenotypically (Figure 1), the inheritance of each is quite different. Normally, sterile brachytic plants result from cross combinations between subspecies; whereas, sterile dwarf can result from crosses within the subspecies, hypogaea.

Shows two dug and inverted sterile brachytic plants (top) and two sterile dwarf plants (bottom) found in F2 populations resulting from cross combination between subspecies hypogaea x fastigiata and within subspecies hypogaea x hypogaea, respectively.
During 2014, only one of three F3:4 families from another cross combination (C1803) involving the backcross [(Georgia-06G X GA 032913) X Georgia-06G] also resulted in a 3 normal to 1 sterile dwarf plant genetic ratio. The F2 (C1803 and C1813) populations were in close field proximity during 2012. Possibly, a natural cross occurred between the partially sterile dwarf male plant of C1813 and a normal C1803 female plant. The resulting F1 normal plant was then selected during 2013. Subsequently, stained pollen from the flower of sterile dwarf plants was indeed observed to be partially normal and fertile. Obviously, the partially fertile pollen characteristics of the sterile dwarf distinguish it from the male sterility reported by Upadhyaya and Nigam (1996), which was completely sterile and controlled by two recessive genes, ms1 and ms2 .
Consequently in crosses involving certain parental lines within the subspecies hypogaea, the homozygous recessive sdw sdw alleles result in approximately 25% sterile dwarf plants in the F2 population, and also two-thirds of the F2 normal plants will be heterozygous (Sdw sdw) which will segregate 2 normal to 1 sterile dwarf plants in the next generation. Without knowing which normal plants are homozygous or heterozygous, individual plant selection can unfortunately perpetuate this undesirable trait into subsequent progeny rows.
Sterile brachytic plants have long been expected from infraspecific crosses (Ashri 1968, Coffelt and Hammons 1972, Hull 1937), and likewise have been observed in later generation progeny from selected normal plants as well. In fact during 2015, a single sterile brachytic plant with tall slender mainstem and flowers was found in an F6 breeding line from the infraspecific cross, C1721 (Georgia-11J X Georgia Valencia).
Molecular markers would be extremely helpful to differentiate individual plant selections carrying these deleterious gene(s) for both sterile dwarf and sterile brachytic plants. Peanut breeders could then utilize marker assisted selection (MAS) to avoid these two dwarf traits in early-generation progeny rows.
Acknowledgements
The authors wish to particularly thank Professor (Emeritus) Amram Ashri at the Hebrew University of Jerusalem, Rehovot, ISRAEL for his very helpful review of the manuscript prior to its submission.
Literature Cited
A Ashri, (1968). Morphology and inheritance of sterile brachytic dwarfs in peanuts, Arachis hypogaea . Crop Sci 8 : 413 – 415 .
W.D Branch, (2001). Registration of ‘Georgia Valencia' peanut. Crop Sci 41 : 2002 – 2003 .
W.D Branch, (2007). Registration of ‘Georgia-06G' peanut. J. Plant Reg 1 : 120 .
W.D Branch, (2010). Registration of ‘Georgia-09B' peanut. J. Plant Reg 4 : 175 – 178 .
W.D Branch, (2012). Registration of ‘Georgia-11J' peanut. J. Plant Reg 6 : 281 – 283 .
T.A Coffelt, and R.O Hammons (1972). Inheritance of sterile brachytic in an infraspecific cross of Arachis hypogaea L. Crop Sci 12 : 82 – 84 .
Hammons, R. O 1973 Genetics of Arachis Hypogaea Pp 135 – 174 In: Peanuts – Culture and Uses, Amer. Peanut Res. Educ. Assn Inc ., Stillwater, OK .
C.C., Holbrook, P Timper, A.K Culbreath, and C.K Kvien (2008). Registration of ‘Tifguard' peanut. J. Plant Reg 2 : 92 – 94 .
Hull, F.H 1937 Inheritance of rest period of seeds and certain other characteristics in the peanut. Univ. Florida Agr Exp. Stn. Tech. Bull. 314 .
J.S., Patel, C.M John, and C.R Seshadri (1936). The inheritance of characters in the groundnut, Arachis hypogaea . Proc. Indian Acad. Sci 3 ((B)) : 214 – 233 .
Strickberger, M. W 1968 Chap. 8. Probability and statistical testing Pp 126 – 152 In: Genetics The Macmillian Co ., New York, NY .
H. D Upadhyaya, and S. N Nigam (1996). Identification and inheritance of male sterility in groundnut ( Arachis hypogaea L.). Euphytica 88 : 227 – 230 .
Notes
- Professor, University of Georgia, Department of Crop & Soil Sciences, Coastal Plain Experiment Station, 2360 Rainwater Road, Tifton, GA 31793-5766. [^]
- Graduate Student, Institute of Plant Breeding, Genetics & Genomics, University of Georgia, Tifton, GA 31793-5766. [^]
- Assoc. Professor, University of Georgia, Department of Crop & Soil Sciences, Coastal Plain Experiment Station, 2360 Rainwater Road, Tifton, GA 31793-5766. [^] *Corresponding author email: wdbranch@uga.edu
Author Affiliations