

Introduction
The nitrogen fixation process in peanut (Arachis hypogaea L.) is essential for providing nitrogen (N) for the crop throughout the season, as it delivers from 50-80% of the total N required by the plant (Boddey et al., 1990). The rates of N fixation for peanut range from 100 to 190 kg N/ha (Boddey et al., 1990), and are quite comparable to other N-fixing crops such as soybean [Glycine max (L.) Merr.; 85 – 155 kg N/ha], chickpea [Cicer arietinum L.; 103 kg N/ha], or pigeon pea [Cajanus cajan (L.) Mills.; 168 – 280 kg N/ha) (Elkan, 1995). The process of biological N fixation (BNF) involves symbiotic bacteria that reduce atmospheric N2 to ammonia, a form of N that is available for plant nutrition (Nievas et al., 2012). The BNF process can account for as much as 65% of the N that is used in agriculture worldwide, with the largest quantity of fixed N originating from the symbiotic association between rhizobia and legumes (Dupont et al., 2012). For example, intercropped peanut has the capacity to provide over 10% of the total N that is accumulated in the accompanying crop during a growing season (Chu et al., 2004). Therefore, assessing the process of nodulation in peanut could have important implications for evaluating and developing sustainable cropping systems globally.
The process of infection of roots by N fixing bacteria is fairly unique in peanut and occurs through a process known as “crack entry” (Boogerd et al., 1997). The process involves the invasion of rhizobia between the epidermal cells at the base of emerging lateral roots, and the eventual proliferation of bacteria into the root cortical cells (Bogino et al., 2011; Nievas et al., 2012). The majority of nodules in peanut are formed on the first order lateral roots near the basal region (top 8 cm) of the taproot (Tajima et al., 2006). Peanut generally lacks root hairs on the taproot because the surface cell layers are shed, so nodules form at the base of lateral roots where rosette-type root hairs up to 4 mm in length are located (Akasaka et al., 1998). Peanut typically forms determinate type nodules (Akasaka et al., 1998) that are characterized by having no permanent meristem, a regular spherical shape, and a homogeneous central tissue within the nodule that is composed of tightly packed bacteroids responsible for the actual N fixation process (Desbrosses and Stougaard, 2011; Dupont et al., 2012). Alternatively, legumes such as pea (Pisum sativum L.) or clover (Trifolium spp.) form indeterminate nodules that have a permanent meristem and take on a more cylindrical shape (Dupont et al., 2012). The two primary bacterial genera that commonly colonize peanut nodules are Bradyrhizobium as well as Rhizobium (Nievas et al., 2012; Angelini et al., 2011). However, rhizobia within single peanut nodules can be very diverse with no real dominant strain (Angelini et al., 2011) and often include non-rhizobial endophytes (Ibáñez et al., 2009).
Aside from biological factors such as rhizobial strain or host specificity among crop cultivars, environmental conditions and management practices within typical field production settings can impact the formation of nodules and the subsequent process of N fixation. These factors include: soil temperature; water availability; soil acidity; soil salinity; intercropping; crop rotation; fertilization regime; inoculation practices; and tillage practices (Sprent, 1972; Hungria and Vargas, 2000; van Kessel and Hartley, 2000; El-Akhal et al., 2013). Reduced tillage and cover crops prior to a legume crop often increase nodulation, as is the case for soybean nodulation (Hughes and Herridge, 1989) and rates of N fixation (Kihara et al., 2011) for this crop. Increased N fixation has also been noted under reduced tillage for chickpea and faba bean (Vicia faba L.; López-Bellido et al., 2011). One typical system where tillage plays a critical role and impacts the performance of the subsequent peanut crop is bahiagrass (Paspalum notatum Flueggé) sod systems. Bahiagrass sod is an important economic component in many Florida peanut production systems either through inclusion of bahiagrass and livestock in rotation with peanut (Katsvairo et al., 2006) or because many producers may rotate long-term bahiagrass fields with peanut to economically benefit their production system. Tillage systems that can be utilized to break up the bahiagrass sod may be limited due to its extensive and fibrous root system. Conventional, deep tillage is primarily chosen, but some growers are considering the economic and environmental benefits of reduced tillage systems (such as strip tillage) that are able to reduce fuel use and protect soil resources. Therefore, the tillage operation chosen to remove the bahiagrass sod could have serious impacts on the nodulation pattern and N fixation of the following peanut crop.
While it is acutely important to assess the impact of tillage and other field management decisions on nodulation and N fixation rates, evaluation and quantification of nodulation patterns in the field is difficult and partially explains the paucity of studies that have successfully achieved these measurements. Methods typically involve the destructive harvest of roots through coring, trenching, or uprooting large quantities of soil – all techniques that are extremely labor intensive, hugely destructive within a research plot area, and most often preclude the ability to perform repeated sampling within a growing season (Gray et al., 2013). Recently however, Gray et al. (2013) successfully demonstrated the efficacy of using the minirhizotron technique for the study of nodulation patterns in field-grown soybean under elevated CO2 combined with drought stress. The minirhizotron technique involves the installation of clear plastic tubes within and parallel to the crop row, usually at a 30 or 40° angle which allows for repeated imaging of the root system through the season (Milchunas, 2012). A camera is inserted within the tube and roots with associated nodules can be imaged along the length of the tube. Images can then be digitally analyzed for nodule number, size, and density as well as other root traits through the use of several software programs. Because the tube normally has a locking mechanism interfaced with the camera, the user can repeatedly image the exact same locations along the length of the tube over time.
The minirhizotron technique may allow for the detailed and comprehensive examination of the natural history of peanut nodule formation by soil depth and across time that has not been previously documented. In addition, the effect of alternative tillage systems on nodulation, particularly those utilized in a bahiagrass sod rotation can also be examined. To investigate the suitability of the minirhizotron imaging technique in determining differences in peanut nodulation, a field experiment was established in Citra, FL using alternative tillage operations in a bahiagrass sod cropping system as a test case. In situ nodulation patterns along the root system over the growing season were imaged directly through the use of minirhizotrons.
Materials and Methods
Tillage Treatments and Crop Maintenance
In 2012, field plots were established at the University of Florida’s Plant Science Research and Education Unit (PSREU) located near Citra, FL (29°24’28” N, 82°10’30” W, elevation 21 meters) on a Sparr fine sand (loamy, siliceous, sub active, hyperthermic Grossarenic Paleudults). Plots were arranged in a randomized complete block design with the following treatment factors: three tillage treatments (Conventional, Strip, and Strip followed by use of a High Residue Cultivator – Strip/HRC), and two timings of tillage (all tillage operations one month prior to planting – Date 1, and all tillage at the time of planting – Date 2). The crop followed a well-established stand of bahiagrass. The entire plot area was killed chemically with a 2% (1.9 L/ha) rate of glyphosate. The vegetation in the field was allowed to die down for one month before the early tillage (Date 1) operations. The conventional plots were disked three times, turned with a moldboard plow and smoothed with a disc harrow and field cultivator. The strip tillage plots were stripped using a KMC Rip Strip unit outfitted with subsoil shanks and wavy coulters (Tifton, Ga). For the Strip/HRC treatment, at approximately 50 days after planting the plots were cultivated using a KMC Hi Residue Cultivator (Tifton, Ga).
After completion of the Date 2 tillage operations, all plots were planted on 9 May with the commercial peanut cultivar, Florida-07 (Gorbet and Tillman, 2009) using a Monosem (Edwardsville, KS) air planter with an in-row seed population of 19.7 seed/m. Plots consisted of 8 rows (91 cm spacing) 30.5 m long. At planting, a granular inoculant (Cell-Tech®, Novozymes) was applied to all plots using a hopper box attached to the planter. Supplemental seed was added by hand to row areas across the field that had inadequate emergence and stand populations less than 13.1 seed/m on 22 May. Management of pesticides and fertilization was conducted according to the University of Florida’s Institute of Food and Agricultural Sciences (IFAS) recommendations (Table 1).
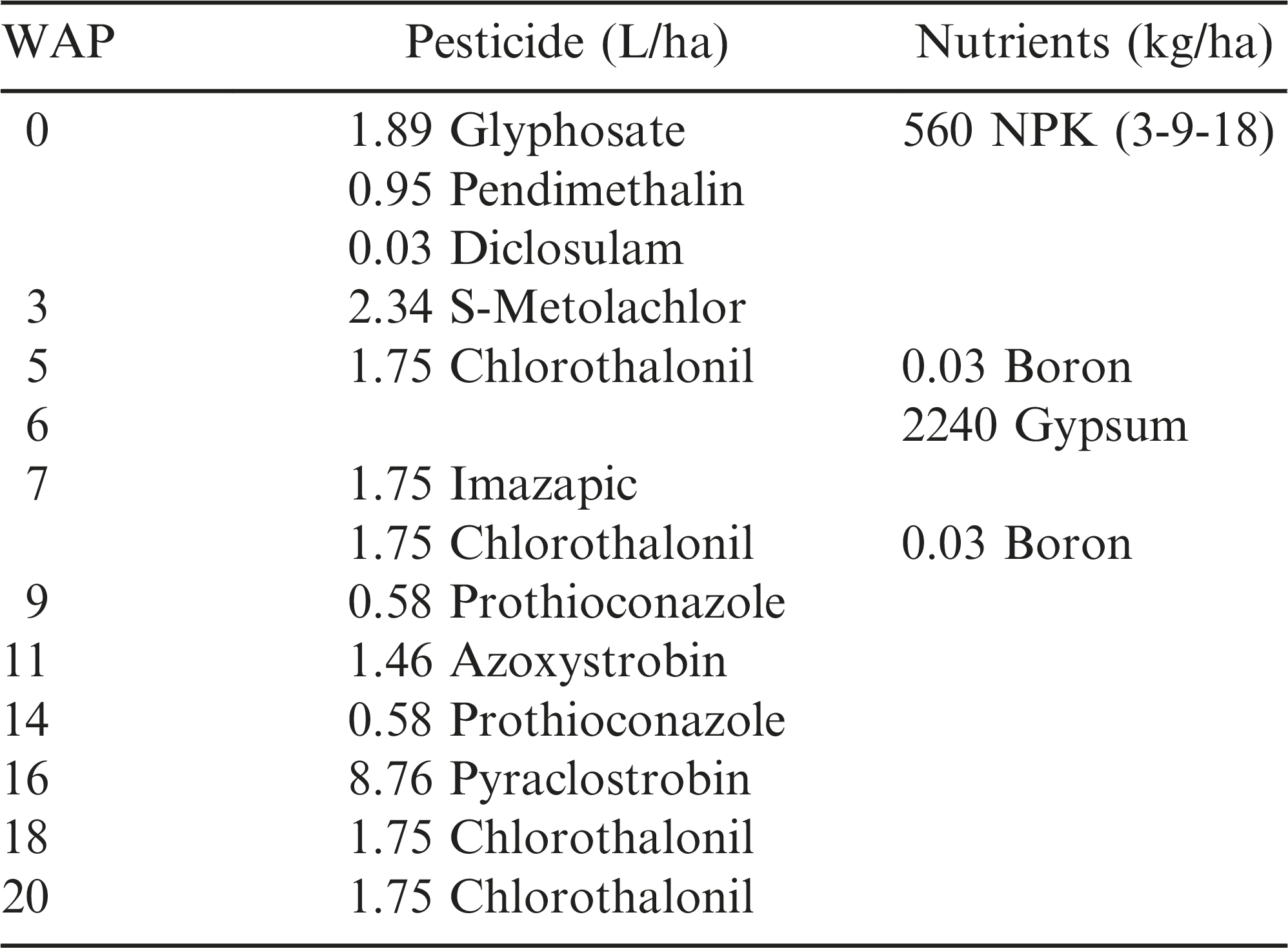
The weeks after planting (WAP) when applications of pesticides and nutrients were made during the experiment at PSREU in 2012.
Measurement of Root System and Nodulation
Nodule establishment and growth were assessed by regularly recording root images through the season in the installed minirhizotron tubes from the soil surface to a depth of approximately 90 cm. To accomplish this, clear plastic minirhizotron tubes (183 cm in length; Bartz Technology, Carpinteria, CA) were inserted in-row and parallel to the crop at a 45° angle with the soil surface in each plot. Roots were digitally imaged on four dates (12 June, 9 July, 1 August, and 17 August) within ordered frames (13 X 18 mm area) along the entire length of the tube using a minirhizotron camera system (Bartz Technology, Carpinteria, CA). The locking mechanism of the camera allowed for repeated imaging of identical locations within the tube at every imaging session. When nodules were present within image frames, depth was calculated and nodules were counted and their size determined using the Image J (National Institutes of Health) software program which calculates the diameter and surface area of each nodule. Nodule density was calculated using the frame area of each image.
Statistical Analysis
Data for total number of nodules and nodule surface area were analyzed using ANOVA in SAS JMP (SAS Institute, Inc., Cary, NC) and Tukey’s HSD multiple comparisons test was employed to determine the separation among mean values in these two variables.
Results
Nodule formation was successfully imaged using the minirhizotron technique, allowing for the sequential measurement of the number and size of nodules through the growing season. It was possible to return to the same imaging frame at each sampling date which allowed the phenological study of individual nodules over time. An interesting observation was noted: many nodules were surrounded by air spaces which may be important for adequate gas exchange that must occur during the N fixation process (Figure 1).

Observations of air spaces surrounding nodules was common in the current study and may indicate an important factor supporting gas exchange during the N fixation process.
Utilizing the minirhizotron system in the bahiagrass sod test case, the pattern of nodulation within the different tillage systems could be assessed with the technique. In this test case, the individual tillage operations had no impact on the total number of nodules at all soil zone depths and image dates (Table 2). However, the date when the tillage operations were performed (approximately 30 days prior to planting – Date 1, or at plant – Date 2) did influence the number of nodules produced, with Date 2 having more nodules than the earlier tillage date. There were no differences in nodule number among the four imaging dates. This indicates that the number of nodules formed was fixed relatively early in crop development, at least by the second image data (9 July, 61 DAP), and stayed constant through the season with few new nodules formed or older nodules deteriorated. This is illustrated in Figures 2 and 3 where groups of nodules became apparent by 9 July (61 DAP) and persisted until the last image date on 17 August (100 DAP). This indicates that nodules are fully formed by the crop’s peak pod fill (60 DAP) and persist through peak physiological performance (90 DAP). There was one instance where nodules were noted to have senesced and disappeared (Figure 4), but based on the analysis results, this was a fairly rare occurrence. Nodule number differed among soil depth zones (Table 2), with the majority of the nodules formed within soil depth zones 1 and 2 (0-20 cm in depth, Figure 5). The deepest single nodule in the study was noted at 90 cm below the soil surface in the Strip/HRC treatment and persisted through to the 17 August imaging date (Figure 6).

Sequential images of a peanut root system illustrating nodule development and possible senescence within the study site taken on: 12 June (a), 9 July (b), 1 August (c), and 17 August (d).

Sequential development of nodules over time on two root segments illustrating nodule establishment and possible senescence. Images taken on 12 June (a,d), 1 August (b,e), and 17 August (c, f); note that the images taken on 9 July were identical in nodule size and color as the images from 1 August, but due to a preferred background color, the 1 August images are shown.

Root segment image illustrating the process of nodule senescence and eventual deterioration: nodules are visible on root segment 9 July (a) but are absent on 1 August (b).
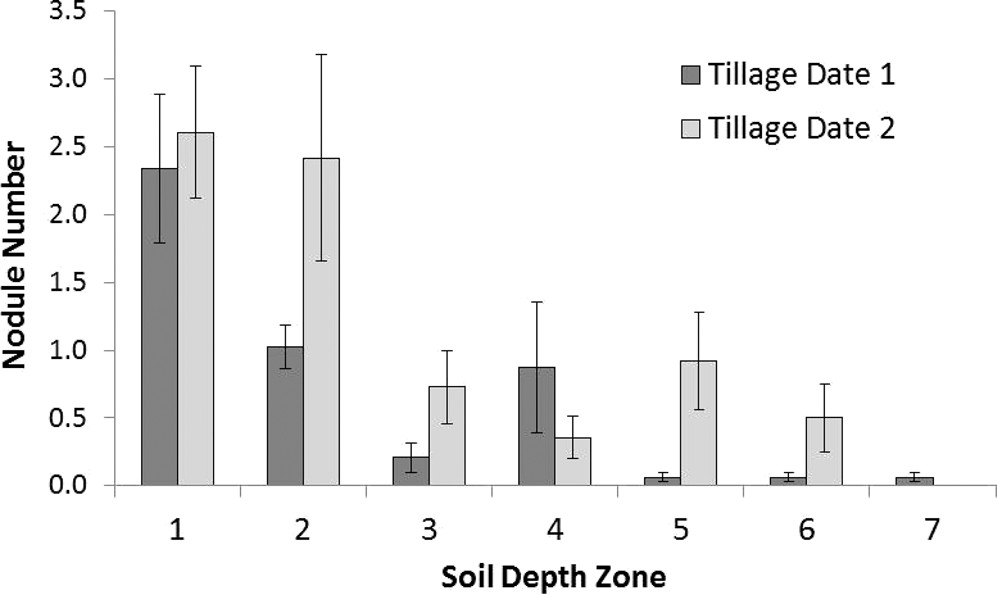
Average number of nodules by soil depth zone across all tillage treatments but separated by the date of the tillage operation (Tillage Date 1 – one month prior to planting; Tillage Date 2 – at planting). Zones are depths from the soil surface: 1 (0-10 cm), 2 (10-20 cm), 3 (20-30 cm), 4 (30-40 cm), 5 (40-50 cm), 6 (50-60 cm), and 7 (60-70 cm).

The deepest single nodule in the 2012 study was located at 90 cm below the soil surface. The nodule was located in the Strip/HRC treatment and persisted until the last imaging date (17 August).
To illustrate general nodule density and the pattern of density development by depth across the primary root area examined, nodule density was calculated as the number of nodules within the reading frame area (13 X 18 mm = 234 mm2 area) and graphed along the rooting depth imaged (roughly 90 cm below the soil surface). Figure 7 illustrates the general pattern of nodule density by soil depth across all tillage and tillage date treatments for three imaging dates. This illustrates how nodules were clustered within the top 20 cm of soil and that the nodule density became fairly constant by 9 July. Nodule density below 40 cm was only noted for 9 July and 17 August, likely reflecting the overall pattern of root extension to deeper soil depths by 9 July.
Nodule size expressed as surface area differed among tillage treatments with Strip and Strip/HRC both having larger nodules than Conventional tillage (Table 2). This was the case regardless of when the tillage operation was performed (Date 1 or 2) resulting in average nodule surface areas of 1.9, 2.7, and 2.5 mm2 for Conventional, Strip, and Strip/HRC, respectively (Figure 8). There were differences among the image dates with the maximum nodule surface area being reached on 1 August; the surface area values at this date were significantly different than the first image date on 12 June. Visually, the expansion of nodule size is illustrated by images in Figure 9 where the enlargement of a single nodule is shown. Nodule surface areas were different among soil depth zones with larger nodules congregated in the top 20 cm (Figure 10). There was an interaction between tillage and depth zone (Table 2) driven by the trend of larger nodules in the Strip/HRC in zone 2 more so than the other two tillage treatments (Figure 10).
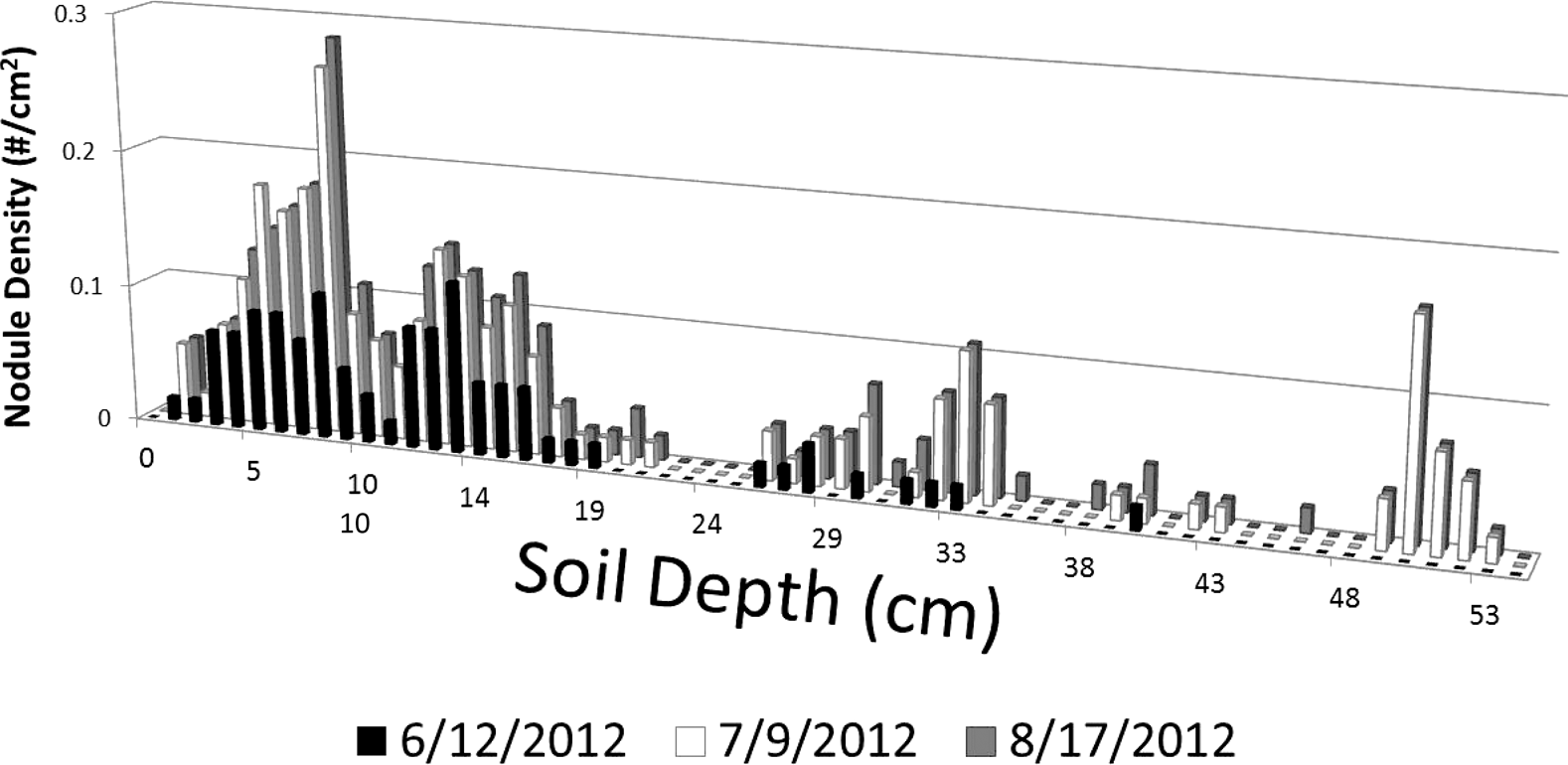
Average nodule density (# nodule per soil area within reading frame – 13 X18 mm) by depth. Patterns of nodule density are shown for three imaging dates 12 June, 9 July, and 17 August.
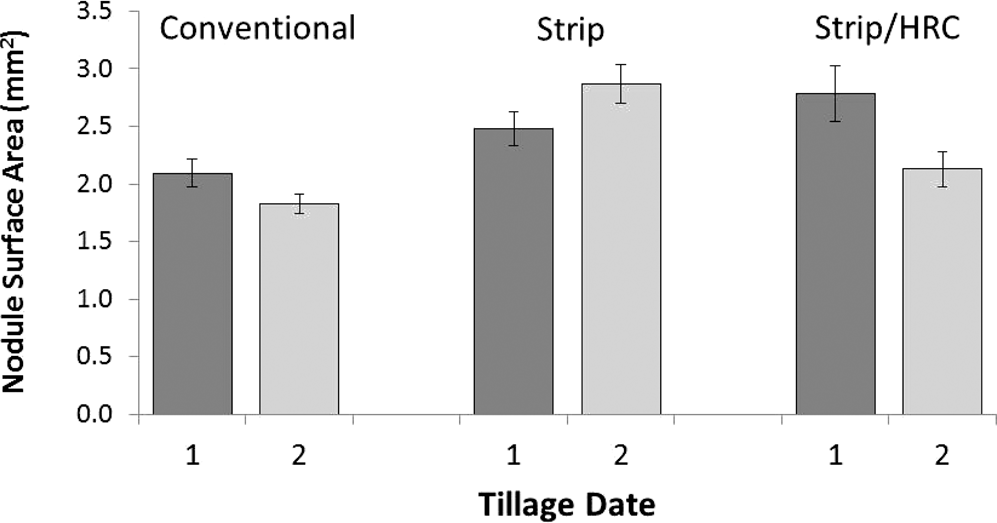
Average individual nodule surface area (mm2) across all soil depths and imaging dates. Results are shown for three tillage treatments (Conventional, Strip, and Strip/HRC) and the dates of tillage operation (1 month prior to planting – Date 1, and at planting – Date 2).

Illustration of nodule increase in girth over time. Images were taken 12 June, 1 August, and 17 August.
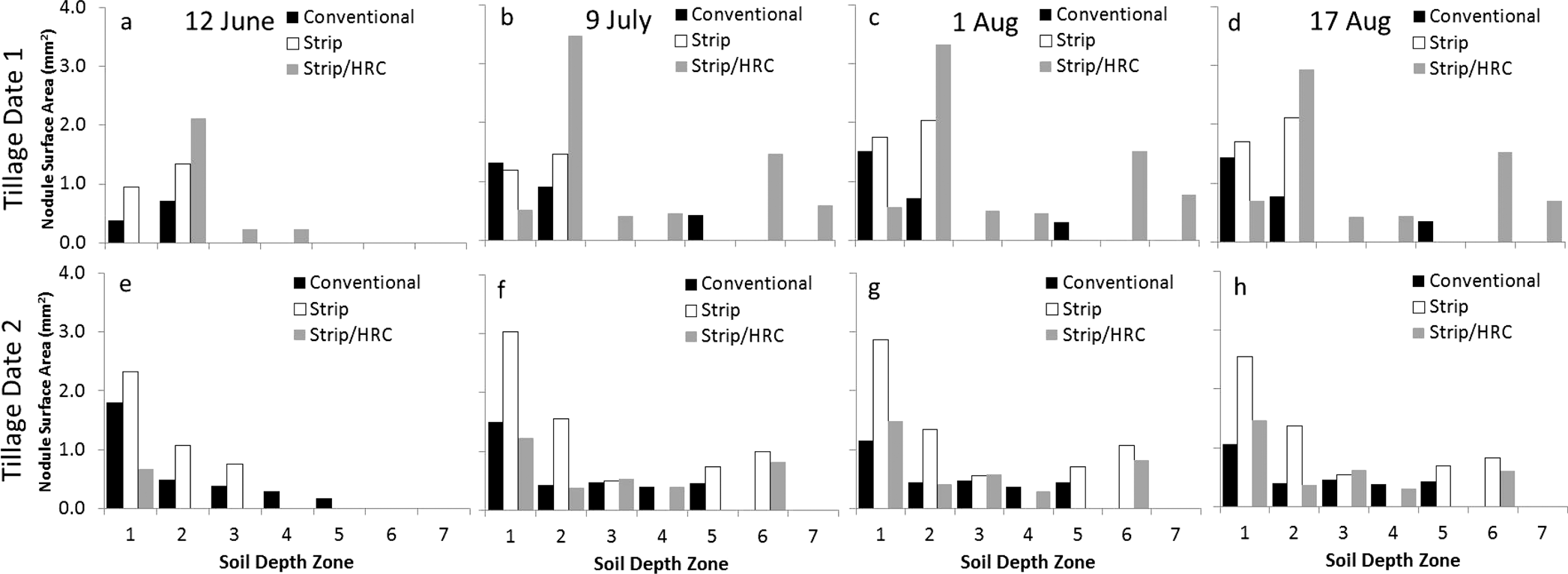
Nodule surface area by soil depth zone. Zones are depths from the soil surface: 1 (0-10 cm), 2 (10-20 cm), 3 (20-30 cm), 4 (30-40 cm), 5 (40-50 cm), 6 (50-60 cm), and 7 (60-70 cm). Treatments include Tillage Date 1 (a-d) and Tillage Date 2 (e-f); within each tillage date, four imaging dates are included: 12 June (a,e), 9 July (b,f), 1 August (c,g) and 17 August (d,h).
Discussion
There are several important contributions from this study: 1) the general observational and quantitative assessment of the natural history and progression of peanut nodule formation, both temporally and spatially; 2) the evaluation of the minirhizotron system in a test case examining the impact of tillage on this natural history; and 3) the demonstration of the distinctive and expansive research applications that the minirhizotron technology can afford when studying belowground processes. In general for most crop species, the pattern of root nodule formation in relation to different soil depth zones has rarely been reported (Tajima et al., 2006). The minirhizotron system is perfectly suited for this type of study, but this technique has not been commonly utilized to date for studying nodulation patterns (but see Gray et al., 2013), and the current study is certainly the first for peanut.
Based on the current results, the nodulation pattern over time indicated that nodules were formed relatively early in the growing season and the number was set by 9 July (61 DAP) corresponding to peak pod fill (Boote, 1982). New nodule formation or nodule senescence past that date was relatively rare. This pattern may be due to the suppressive ability of early-formed nodules on subsequent nodule development (Kossiak and Bohlool, 1984; Danso and Bowen, 1989). This timing of nodule production for peanut is similar to studies on other species noting that primary nodulation is attained by crop flowering (Voisin et al., 2003). As far as nodule pattern within the root system, the current study showed most nodules were formed within the top 20 cm below the soil surface, a pattern that is also similar to other studies (Danso and Bowen, 1989; Hardarson et al., 1989; Tajima et al., 2006). However, the current study did note an increase in nodule surface area over time indicating that, while nodule number may have been set early in crop development, nodule expansion was certainly possible through the season.
The variability among different soil depth zones in the nodule density and size found in this study has important implications regarding the rhizobial population that was likely infecting the different root segments. Native rhizobial populations are normally widely distributed throughout the soil profile (Bogino et al., 2011). This is unlike the patterns of distribution for rhizobial strains used in commercial inoculants applied in-furrow or directly on the seed that normally remain near the soil surface (0-10 cm); this is because movement of these rhizobial strains through the soil profile is very slow (Hardarson et al., 1989; McDermott and Graham, 1989; Bogino et al., 2011). Native rhizobial populations are also highly competitive against commercial strains and will normally interfere with the effectiveness of infection by these applied inoculants (Bogino et al., 2011). In the current study, the zones where nodule density and size were high occurred in two zones, 0-10 cm and 10-20 cm. This indicates that perhaps the applied inoculant in the 0-10 cm and native rhizobial strains in the 10-20 cm and deeper soil zones were both active and effective. This is strictly speculative, but testing to confirm and quantify the contribution of native rhizobia seems important in evaluating the effectiveness of commercially applied inoculant. The soil depth zone position of nodules is also important because it is known to directly impact N fixation activity (Hardarson et al., 1989). Roots near the surface derive a higher proportion of their N from the symbiosis with rhizobia (Lory et al., 1992) while deeper nodules have relatively low levels of N-fixation (McDermott and Graham, 1989; Tajima et al., 2006), sometimes due to inhibitory levels of incorporated N fertilizer. However, conservation tillage systems that provide limited soil mixing may have lower nutrient levels at deeper depths which may actually help increase the N-fixing activity of nodules within these regions because the nutrient concentrations would rarely reach inhibitory levels (Virginia et al., 1986).
Nodule size is known to have a direct impact on N fixation (Voisin et al., 2003; Tajima et al., 2007) particularly under stress conditions (King and Purcell, 2001). Large nodules may have amplified N fixation activity in comparison to small nodules due to a greater phloem supply (Purcell et al., 1997). It has been estimated for soybean that, by volume, 25% of a 2 mm diameter nodule is infected, while a 4 mm diameter nodule has 60% infected tissue indicating greater relative N fixation ability (King and Purcell, 2001). In general, peanut has smaller nodules than soybean, but higher overall N fixing activity (Tajima et al., 2007). This size range was similar to that found in the current study where most peanut nodules were approximately 2 mm2 in surface area, translating to a 1.4 mm diameter.
This case study was successful in evaluating the efficacy of the minirhizotron system for evaluating the impact of tillage practices on the natural history of nodulation including responses in nodule number and size. Previous studies have found a promotion of nodulation under no-tillage systems and that there was a close positive relationship between nodule number and the N-content of roots within conservation tillage systems (Hughes and Herridge, 1989; Sidiras et al., 1999; Kihara et al., 2011; López-Bellido et al., 2011). Tillage system in the current case study had no effect on nodule number but did significantly influence nodule surface area (a surrogate for overall nodule size). The current results indicate that nodule surface area was enhanced in both conservation tillage treatments (Strip and Strip/HRC), regardless of whether the tillage operation was performed one month prior or at planting. Because of the strong relationship between nodule size and N-fixation rates (Purcell et al., 1997; King and Purcell, 2001; Voisin et al., 2003; Tajima et al., 2007), this result indicates a possible advantage of conservation tillage systems through enhancement of N-fixation rates as a whole. However, because the data was relatively limited in scope and primarily aimed at evaluating the use of the minirhizotron technique in different tillage systems, these results require further study and confirmation.
Another contribution from this study is the illustration of the application of the minirhizotron technique in a myriad of possible studies focused on belowground processes. Aside from root measurements and nodulation patterns, the minirhizotron images captured in this study illustrate additional assessments of belowground community dynamics that could be possible using the technique. Other soil organisms were captured in the images from the current study including arthropod egg masses (Figure 11). Studies concentrating on populations of soil organisms could also be digitally imaged using this technique. In the particular case study, further examination of the interaction between the endophytic fungi of bahiagrass and rhizobia on subsequent colonization in peanut could also be studied using minirhizotrons. The current study and similar applications of the minirhizotron technique show the potential of this method to provide novel and important imaging of phenomena that have previously been unobservable. This was certainly the case for the current study’s documentation of nodulation patterns and processes and opens the door for additional unique investigations.

Image of nodule (a) and possible arthropod egg masses (b) within a soil cavity; image taken on 12 June.

ANOVA results for total number of nodules and nodule surface area as influenced by image date (12 June, 9 July, 1 August, and 17 August), tillage (Conventional, Strip Till, Strip/HRC), tillage date (Date 1, 2), and soil depth zone (0-10, 10-20, 20-30, 30-40, 40-50, 50-60, and 60-70 cm).
Acknowledgements
We thank the staff at the UF Plant Science Research and Education Unit for help with plot and crop maintenance.
Literature Cited
Akasaka Y Mii M and Daimon H 1998 Morphological alternations and root nodule formation in Agrobacterium rhizogenes-mediated transgenic hairy roots of peanut (Arachis hypogaea L.) Ann. of Botany 81 : 355 – 362 .
Angelini J Ibáñez F Taurian T Tonelli M.L Valetti L and Fabra A 2011 A study on the prevalence of bacteria that occupy nodules within single peanut plants Curr. Microbiol 62 : 1752 – 1759 .
Boddey R.M Urquiaga S Neves M.C.P Suhet A.R and Peres J.R 1990 Quantification of the contribution of N2-fixation to field grown grain legumes – a strategy for the practical application of the N15 isotope-dilution technique Soil Biol. Biochem 22 : 649 – 655 .
Bogino Nievas P.F Banchio E and Giordano W 2011 Increased competitiveness and efficiency of biological N fixation in peanut via in-furrow inoculation of rhizobia European J. Soil Biol 47 : 188 – 193 .
Boogerd F.C and van Rossum D 1997 Nodulation of groundnut by Bradyrhizobium: a simple infection process by crack entry FEMS Microbiol. Rev 21 : 5 – 27 .
Boote K.J 1982 Growth stages of peanut (Arachis hypogaea L.) Peanut Sci 9 : 35 – 40 .
Chu G.X Shen Q.R and Cao J.L 2004 Nitrogen fixation and N transfer from peanut to rice cultivated in aerobic soil in an intercropping system and its effect on soil N fertility Plant and Soil 263 : 17 – 27 .
Danso S.K.A and Bowen G.D 1989 Methods of inoculation and how they influence nodulation pattern and nitrogen fixation using two contrasting strains of Bradyrhizobium japonicum Soil Biol. Biochem 21 : 1053 – 1058 .
Desbrosses G.J and Stougaard J 2011 Root nodulation: a paradigm for how plant-microbe symbiosis influences host developmental pathways Cell Host & Microbe 10 : 348 – 358 .
Dupont L Alloing G Pierre O El Msehli S Hopkins J Hérouart D and Frendo P 2012 The legume root nodule: from symbiotic nitrogen fixation to senescence, Pages 137-168 in Nagata T (ed.), Senescence, Intech, Croatia 850 pp.
El-Akhal M.R Rincón A Coba de la Peña T Lucas M.M El Mourabit N Barrijal S and Pueyo J.J 2013 Effects of salt stress and rhizobial inoculation on growth and nitrogen fixation of three peanut cultivars Plant Bio 15 : 415 – 421 .
Elkan G.H 1995 Biological fixation in peanuts, Pages 286-300 in Pattee H.E and Stalker H.T (eds.). Advances in Peanut Science. American Peanut Research and Education Soc. , Stillwater, OK 614 pp.
Gorbet D.W and Tillman B.L 2009 Registration of ‘Florida-07’ Peanut J. Plant Reg 3 : 14 – 18 .
Gray S.B Strellner R.S Puthuval K.K Ng C Shulman R.E Siebers M.H Rogers A and Leakey A.D.B 2013 Minirhizotron imaging reveals that nodulation of field-grown soybean is enhanced by free-air CO2 enrichment only when combined with drought stress Functional Plant Bio 40 : 137 – 147 .
Hardarson G Golbs M and Danso S.K.A 1989 Nitrogen fixation in soybean (Glycine max L. Merrill) as affected by nodulation patterns Soil Biol. Biochem 21 : 783 – 787 .
Hughes R.M and Herridge D.F 1989 Effect of tillage on yield, nodulation and nitrogen fixation of soybean in far north-coastal New South Wales Aust. J. Exp. Agric 29 : 671 – 677 .
Hungria M and Vargas M.A.T 2000 Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis in Brazil Field Crops Research 65 : 151 – 164 .
Ibáñez F Angelini J Taurian T Tonelli M.L and Fabra A 2009 Endophytic occupation of peanut root nodules by opportunistic Gammaproteobacteria Systematic and App. Microbiology 32 : 49 – 55 .
Katsvairo T.W Wright D.L Marois J.J Hartzog D.L Rich J.R and Wiatrak P.J 2006 Sod-livestock integration into the peanut-cotton rotation Agronomy J 98 : 1156 – 1171 .
Kihara J Martius C Bationo A and Vlek P.L.G 2011 Effects of tillage and crop residue application on soybean nitrogen fixation in a tropical Ferralsol Agriculture 1 : 22 – 37 .
King C.A and Purcell L.C 2001 Soybean nodule size and relationship to nitrogen fixation response to water deficit Crop Sci 41 : 1099 – 1107 .
Kossiak R.M and Bohlool B.B 1984 Suppression of nodule development on one side of a split-root system of soybean caused by prior inoculation of the other side Plant Phys 75 : 125 – 130 .
López-Bellido R.J López-Bellido L Benítez-Vega J Muñoz-Romero V López-Bellido F.J and Redondo R 2011 Chickpea and faba bean nitrogen fixation in a Mediterranean rainfed Vertisol: effect of the tillage system European J. Agronomy 34 : 222 – 230 .
Lory J.A Russelle M.P and Heichel G.H 1992 Quantification of symbiotically fixed nitrogen in soil surrounding alfalfa roots and nodules Agron. J 84 : 1033 – 1040 .
McDermott T.R and Graham P.H 1989 Bradyrhizobium japonicum inoculant mobility, nodule occupancy, and acetylene reduction in soybean root system Appl. Environ. Microbiol 55 : 2493 – 2498 .
Milchunas D.G 2012 Biases and errors associated with different root production methods and their effects on field estimates of belowground net primary production, Pages 303-340 in Mancuso S (ed.), Measuring Roots, Springer-Verlag, Berlin Heidelberg 382 pp.
Nievas F Bogino P Nocelli N and Giordano W 2012 Genotypic analysis of isolated peanut-nodulating rhizobial strains reveals differences among populations obtained from soils with different cropping histories App. Soil Ecology 53 : 74 – 82 .
Purcell L.C deSilva M King C.A and Kim W.H 1997 Biomass accumulation and allocation in soybean associated with genotypic differences in tolerance of nitrogen fixation to water deficits Plant and Soil 196 : 101 – 113 .
Sidiras N Avgoulas C Bilalis D and Tsourgrianis N 1999 Effects of tillage and fertilization on biomass, roots, N-accumulation and nodule bacteria of vetch (Vicia sativa cv. Alexander) J. Agr. & Crop Sci 182 : 209 – 216 .
Sprent J.I 1972 The effects of water stress on nitrogen-fixing root nodules New Phytol 71 : 603 – 611 .
Tajima Lee R.O.N Abe J Lux A and Morita S 2007 Nitrogen-fixing activity of root nodules in relation to their size in peanut (Arachis hypogaea L.) Plant Prod. Sci 10 : 423 – 429 .
Tajima Morita R.S and Abe J 2006 Distribution pattern of root nodules in relation to root architecture in two leading cultivars of peanut (Arachis hypogaea L.) in Japan Plant Prod. Sci 9 : 249 – 255 .
van Kessel C and Hartley C 2000 Agricultural management of grain legumes: has it led to an increase in nitrogen fixation? Field Crops Research 65 : 165 – 181 .
Virginia R.A Jenkins M.B and Jarrell W.M 1986 Depth of root symbiont occurrence in soil Biol. Fert. Soils 2 : 127 – 130 .
Voisin A.S Salon C Jeudy C and Warembourg F.R 2003 Symbiotic N2 fixation activity in relation to C economy of Pisum sativum L. as a function of plant phenology J. Exp. Bot 54 : 2733 – 2744 .
Notes
- Professor, Undergraduate Student, Graduate Student, Graduate Student, Biological Research Associate, and Professor, respectively, Agronomy Department, University of Florida, 3105 McCarty Hall B, Gainesville, FL 32611. *Corresponding author's e-mail: dlrowland@ufl.edu.
Author Affiliations