

Introduction
Peanut (Arachis hypogaea L.) is the fourth major oilseed crop grown worldwide after soybean [Glycine max (L.) Merr.], rapeseed (Brassica napus L.) and cottonseed (Gossypium hirsutum L.) (USDA FAS 2012) and represents a major source of protein and oil in the human diet in developing countries. In 2011, peanut was grown on 21.7 million ha with a total production of 38.6 MMT worldwide (FAOSTAT 2013). With 444,190 ha and 1.65 MMT of production in 2011 (FAOSTAT 2013), the United States was the fourth largest producer of peanut after China, India, and Nigeria. The major peanut producing states in the US are Georgia, Alabama, Texas, Florida, North Carolina and South Carolina. It is also important to note that most of the peanut production in the world is in arid or semi-arid climates, and peanut therefore is rainfed rather than irrigated.
Virginia-type peanut is an important crop for the Virginia-Carolina (VC) region comprising Virginia, North Carolina, and South Carolina. It is a “cash” crop grown annually on over 50,000 to 76,000 ha in all three states bringing important revenues to growers. Historically, with the incident average precipitation of 784 mm during the growing season in this region, the peanut production of over 250,000 tons per year can be easily achieved without supplemental irrigation. This is also why over 85% of the current peanut farming in the VC region is rainfed rather than irrigated. However, depending on soil type and geography, parts of the VC region have experienced inadequate rainfall and temperature extremes during the growing season in some years. In addition, fine sandy loam soils that are predominantly used for peanut production have poor water holding capacity, exacerbating water deficit stress. For example, in 2009, in general a rainy year with mean monthly rainfall of 103 mm, reduced precipitation during June (87 mm) reduced plant growth and resulted in decreased yields (Balota et al., 2012a). Similarly, due to high temperatures and significantly reduced precipitation in 2010, yield and kernel quality were drastically reduced in the VC region (Balota et al., 2012a). Adoption of irrigation is unlikely due to already high peanut production costs in Virginia and the Carolinas and limited water supply. Therefore, increasing yield under drought and heat in the VC region can only be achieved by the development of drought and heat tolerant cultivars.
Peanut crop can be susceptible to drought and high temperature stress (Bell et al., 1994). Temperature extremes and drought stress individually or combined negatively influence the physiological fitness of plants, pod and seed quality, and yield. As such, there is incentive for development of cultivars with superior drought tolerance. This goal can be achieved by understanding how peanut plants successfully cope with drought stress, or alternatively; how some genotypes more successfully cope with drought stress than others.
In recent years, several physiology-based tools were developed to measure the relative levels of stress experienced by plants under drought stress. These tools are valuable for screening germplasm for drought stress tolerance. Some of the most commonly used screening tools include measuring chlorophyll content, specific leaf area (SLA), and canopy temperature depression (Reynolds et al., 2009; Upadhyaya, 2005).
Water use efficiency (WUE) is the total biomass produced per unit of water transpired, and greater water use efficiency in crops is often associated with good adaptation to water stress. Specific leaf area and leaf chlorophyll content measured by soil plant analysis development (SPAD) chlorophyll meter were reported as good surrogates to select peanut genotypes with improved water use efficiency. Significant correlations were also reported among SPAD chlorophyll reading, specific leaf area and specific leaf nitrogen in peanut (Nageswara Rao et al., 2001). Sheshshayee et al. (2006) reported a strong and positive relationship between SPAD chlorophyll reading and WUE. Recent studies have shown that genetic variation for SPAD chlorophyll reading exists in peanut mini-core entries (Upadhyaya, 2005). Because of the correlation between chlorophyll and N content, SPAD chlorophyll readings alone can be used to estimate nitrogen content in several economically important crops (Chapman and Barreto, 1997, Rorie et al., 2011). As such, SPAD chlorophyll reading was recommended as an appropriate selection criterion for screening for improved drought tolerance in peanut because of its high heritability and simplicity (Songsri et al., 2009; Upadhyaya, 2005).
Plants employ various physiological mechanisms to avoid stress. Drought-tolerant cultivars tend to maintain cooler canopies during stress periods (Ayeneh et al., 2002). Canopy temperature measured by infrared thermometers above the crop canopies is gaining wide acceptance as a potential selection tool for improved drought tolerance and crop yields (Balota et al., 2008; Blum et al., 1988; O'Toole et al., 1984). The canopy temperature depression (CTD) method was described as a rapid, non-destructive, and non-disruptive method of germplasm screening for drought stress and yield ( Balota et al., 2012; O'Toole et al., 1984).
Traits like CTD, SPAD chlorophyll reading, and SLA were successfully used as surrogates to identify drought and heat tolerant cultivars in other peanut growing areas of the world. However, very limited research in relation to drought of the virginia-type peanut is available in the sub-humid VC-region. This study aimed to evaluate the suitability of these physiological traits as surrogates for selection of virginia-type peanut cultivars and breeding lines under rainfed and irrigated field trials in the sub-humid VC-region of the United States.
Materials and Methods
Plant Materials and Growth Conditions
Eight peanut lines including commercial cultivars Bailey (Isleib et al., 2011), CHAMPS (Mozingo et al., 2006), and Phillips (Isleib et al., 2006) and advanced breeding lines N04074FCT, N05006, N05008, N05024J, and SPT 06-07 were selected from the Peanut Variety and Quality Evaluation (PVQE) program based on the variability for CTD, SLA, and SPAD chlorophyll content under drought conditions (Balota et al., 2012a, 2012b).
Field experiments were conducted at the Tidewater Agric. Res. and Extension Ctr., Suffolk, VA (36°68′N, 76°77′W, 25 m elevation) in 2011 and 2012 under rainfed and irrigated conditions. The experiment was a split-split plot design with whole plots arranged in randomized complete blocks (RCB). Water regime was the whole plot factor, genotype was the sub-plot factor, and growth stage a repeating measure factor. Measurements were taken on a fully expanded third youngest leaf arising from the main stem at three peanut growth stages, beginning flower (R1), beginning pod growth (R3), and beginning seed growth (R5) (Boote, 1982). Canopy temperature depression was not measured at stage R1 in 2011 and R3 in 2012. Individual plots of 22 m2 (12.2 m each row and 0.91 m between rows) and with approximately 9 seeds m−1 of row were treated as the experimental unit. Cultural practices were performed according to the Virginia Peanut Production Guide (http://www.ext.vt.edu). Air temperature, RH, rainfall, and photosynthetically active radiation (PAR) were continuously monitored next to the plots with a weather station (WatchDog Temperature/RH Station, Model 2450, Spectrum Technologies Inc., Plainfield, IL), in both years. In 2011, 25.4 mm of irrigation was supplied to the irrigated plots on 24 June, 31.8 mm on 1 July, and 22.9 mm on 19 July using a lateral pull boom cart sprinkler irrigation system (E1025 Reel Rain, Amadas Ind., Suffolk, VA). In 2012, plots in the irrigated water regime received 16.5 mm irrigation on 28 June and 15.8 mm on 18 July. Pod yield data were collected at the physiological maturity in both years.
Canopy Temperature Depression
The CTD of each plot was measured with a hand-held infra-red thermometer (IRT) (Agri-Therm Model 6110L, Everest Interscience Inc., Tucson, AZ) twice at the center of each row in two rows, a total of four readings per plot, at approximately 50 cm above the canopy. The CTD was measured as the difference between canopy temperature (Tc ) and air temperature (Ta ) and converted to positive values when canopies were cooler than the air by multiplying the Agri-Therm readings −1.0, i.e. with the formula:
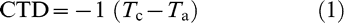
Specific Leaf Area and SPAD Chlorophyll Reading
Four fully expanded third youngest leaves from the main stem per plot were collected in the field and transported to laboratory in moistened paper towels for measurements of leaf area using a LI-3000 leaf area meter (LICOR Inc., Lincoln, NE) and for relative chlorophyll content by the SPAD (Soil Plant Analysis Development) chlorophyll reading (SPAD-502, Konica Minolta Optics Inc., Japan). SLA was calculated after weighing the dried leaves in an oven at 65°C for 24 hours using the formula:
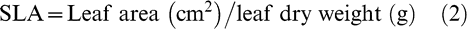
Statistical Analysis
Individual plots were treated as the experimental unit and measurements were taken at three peanut growth stages at beginning flower (R1), beginning pod (R3), and beginning seed (R5). Data from CTD, SLA, and SPAD chlorophyll reading were analyzed as a repeated measures design in a split-split plot arrangement, with growth stage as a repeated measure factor. The univariate model for the repeated measures data was:
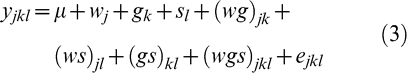
where μ is the overall mean effect, wj the main effect of the j th water regime (j = 1, 2), gk the main effect of the k th genotype (k = 1 to 8), sl the growth stage effect (l = 1 to 3), (wg)jk the interaction of the j th water regime and k th genotype, (ws)jl the interaction of the j th water regime and l th growth stage, (gs)kl the interaction of the k th genotype and l th growth stage, (wgs)jkl the interaction of the j th water regime, k th genotype, and l th growth stage, and ejkl the random error associated with the ijkl th experimental unit. Genotypes and water regimes were treated as fixed effects while the effects of repeated measures, i.e., growth stage, was considered to be random. The sphericity requirements of a univariate repeated measures analysis were tested and then adjusted accordingly using Greenhouse-Geisser (G-G) and Huynh-Feldt (H-F) corrections (Collaku and Harrison, 2002). The data were analyzed using MANOVA option for repeated measures on JMP software (SAS Institute Inc., Ver. 10.0, Cary, NC, 2012).
Correlations between physiological and agronomic traits were calculated. Pod yield was measured at the end of the season and the data analyzed as a split plot design. All the treatments were considered to be fixed in the ANOVA model:
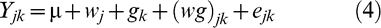
where μ is the overall mean effect, wj the main effect of the j th water regime (j = 1, 2), gk the main effect of the k th genotype (k = 1 to 8), (wg)jk the interaction of the j th water regime and k th genotype, and ejk the random error associated with the experimental unit.
For all traits, mean differences for water regimes and genotypes within a year and growth stage were calculated by Student's t-test (p≤0.05) and Tukey's HSD test (p≤0.05).
Results
Years 2011 and 2012 in general and sampling times (June through August each year) in particular were different from each other in terms of temperature, rainfall, relative humidity (RH), and photosynthetically active radiation (Table 1). Overall, the 2011 growing season was hot, dry with high maximum temperatures (34.1 C), and relatively low precipitation (271 mm) from June to August. Visible symptoms of water deficit stress, i.e., leaf wilting, were observed at stages R1 and R3 in 2011. Later in the growing season, a record of 346 mm rainfall occurred on 27 August, and with repeated rainfall occurring in subsequent days. The 2012 growing season was relatively warm and humid with mean maximum temperatures around 32 C, total rainfall 312 mm, and mean RH of 79% during the June through August interval. During this time, in June specifically, rainfed plants produced less biomass than irrigated plants based on visual comparison, but they were not wilted. From July through October, plots received weekly precipitation, with total rainfall from May to October of 965 mm, approximately 60% more than the multi-annual historical precipitation. Due to persistent cloud cover throughout the entire growing season, photosynthetically active radiation was 30% less in 2012 than in 2011.
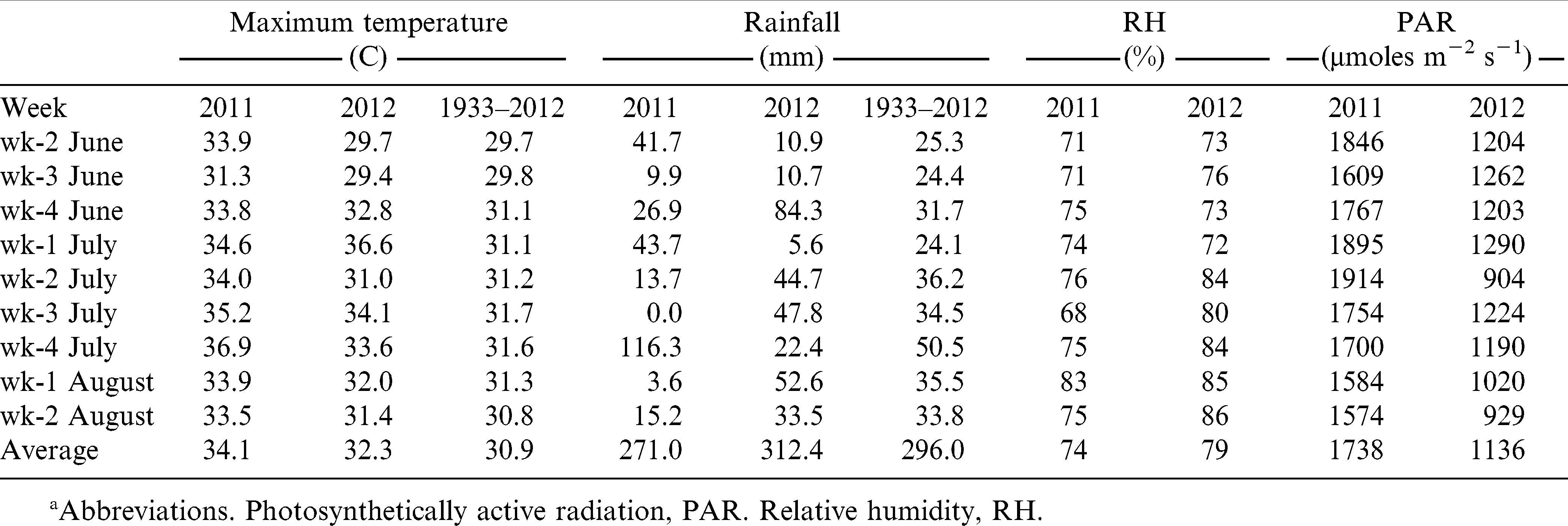
Maximum temperature, total rainfall, relative humidity, and photosynthetically active radiation during June–August in Suffolk, VA in 2011 and 2012. For comparisons, long-term (1933–2012) maximum temperature and rainfall are also presented.
Pod Yield
ANOVA for pod yield showed significant main effects of water regime and genotype in both years, and a significant water regime-by-genotype interaction effect in 2012 (Table 2). In 2011, rainfed plots yielded less than irrigated plots, which received an additional 80-mm water through irrigation during June and July (5783 vs. 6128 kg ha−1, p≤0.05). In 2012, average rainfed plots were higher in yield than irrigated plots (4359 vs. 4185 kg ha−1, p≤0.05) probably due to reduced radiation and excessive moisture on the irrigated plots during the latter part of the 2012 season. N04074FCT had the lowest average pod yield and N05008, N05024J, and Phillips the highest among the genotypes tested under rainfed conditions during both years (Table 3). N05006 had ranked third greatest for yield in 2012, but performed poorly in 2011, the drier year. Low yields under rainfed conditions were recorded for CHAMPS, Bailey, and SPT 06-07 in both years. N05008 had the highest rainfed yield in 2011, but its yield reduction was 8% as compared to its irrigated yield (6360 vs. 7147 kg ha−1). Bailey, CHAMPS, N04074FCT and SPT 06-07 had the greatest reduction of average yield (6.7 to 7.9%), and Phillips and N05006 the lowest (3.9 and 5.5%, respectively) due to water deficit in 2011. In 2012, irrigated SPT 06-07 had a reduction of 11.2% in pod yield compared with its rainfed yield. For all other genotypes yield was higher in rainfed than in irrigated plots in 2012 (Table 3).
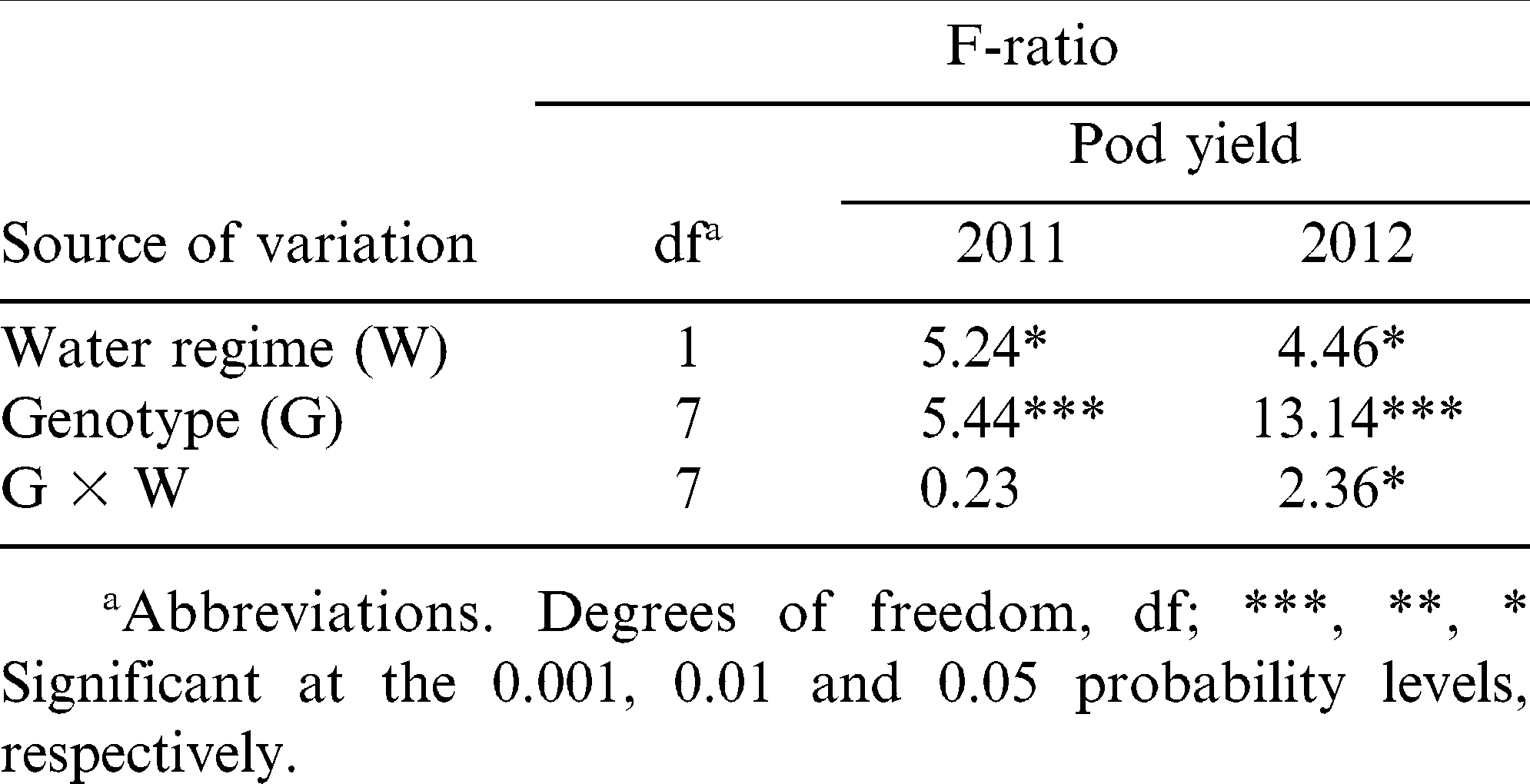
Analysis of variance for pod yield of eight virginia-type peanut genotypes under two water regimes (rainfed and irrigated) in 2011 and 2012.
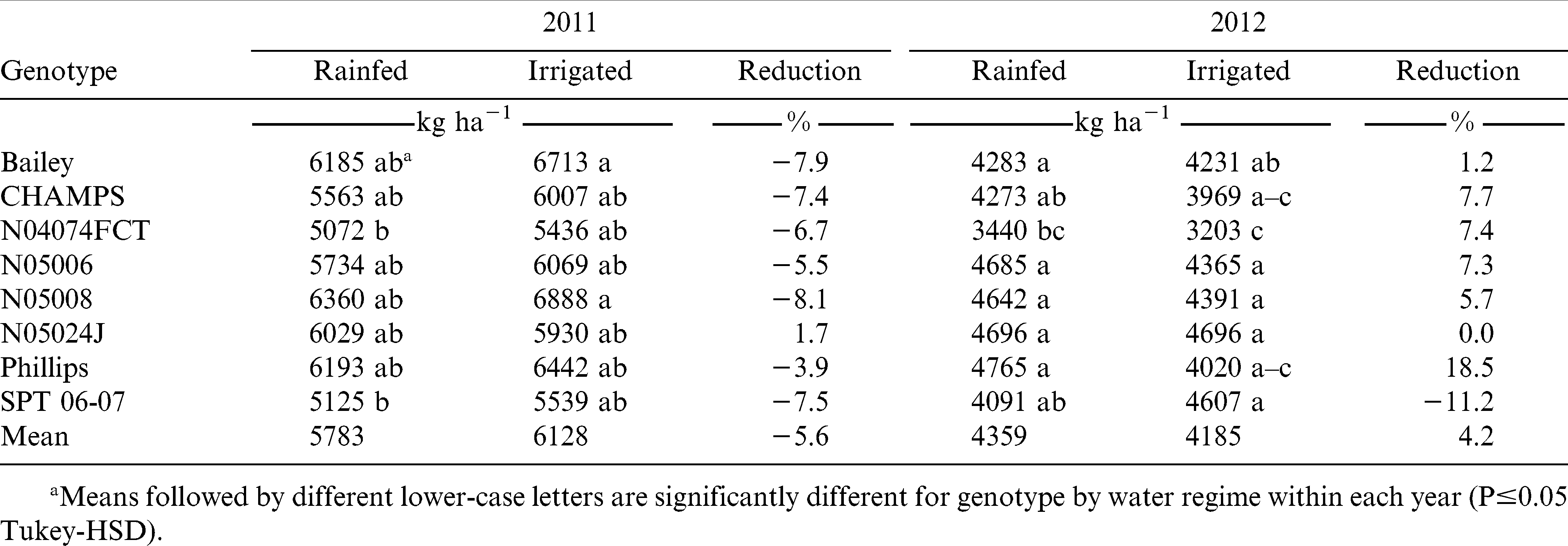
Pod yield and reduction of eight virginia-type peanut genotypes under two water regimes (rainfed and irrigated) in 2011 and 2012.
Canopy Temperature Depression
Univariate repeated measures ANOVA of CTD showed significant water regime and growth stage-by-water regime interaction effects in 2011 and 2012 (Table 4). Growth stage effects were highly significant in 2011 (p≤0.001) and were absent in 2012. In 2011, irrigated and rainfed plants were similarly cooler relative to air temperature (2.4°C) at stage R3, but irrigated plants became significantly cooler than rainfed plants later in the season, at stage R5 (p≤0.05) (Table 5). In 2012, rainfed plots were on average 1.7°C cooler than irrigated canopies during R1 and R5 growth stages. Within each growth stage and water regime, there were no significant differences for CTD among genotypes in both years.
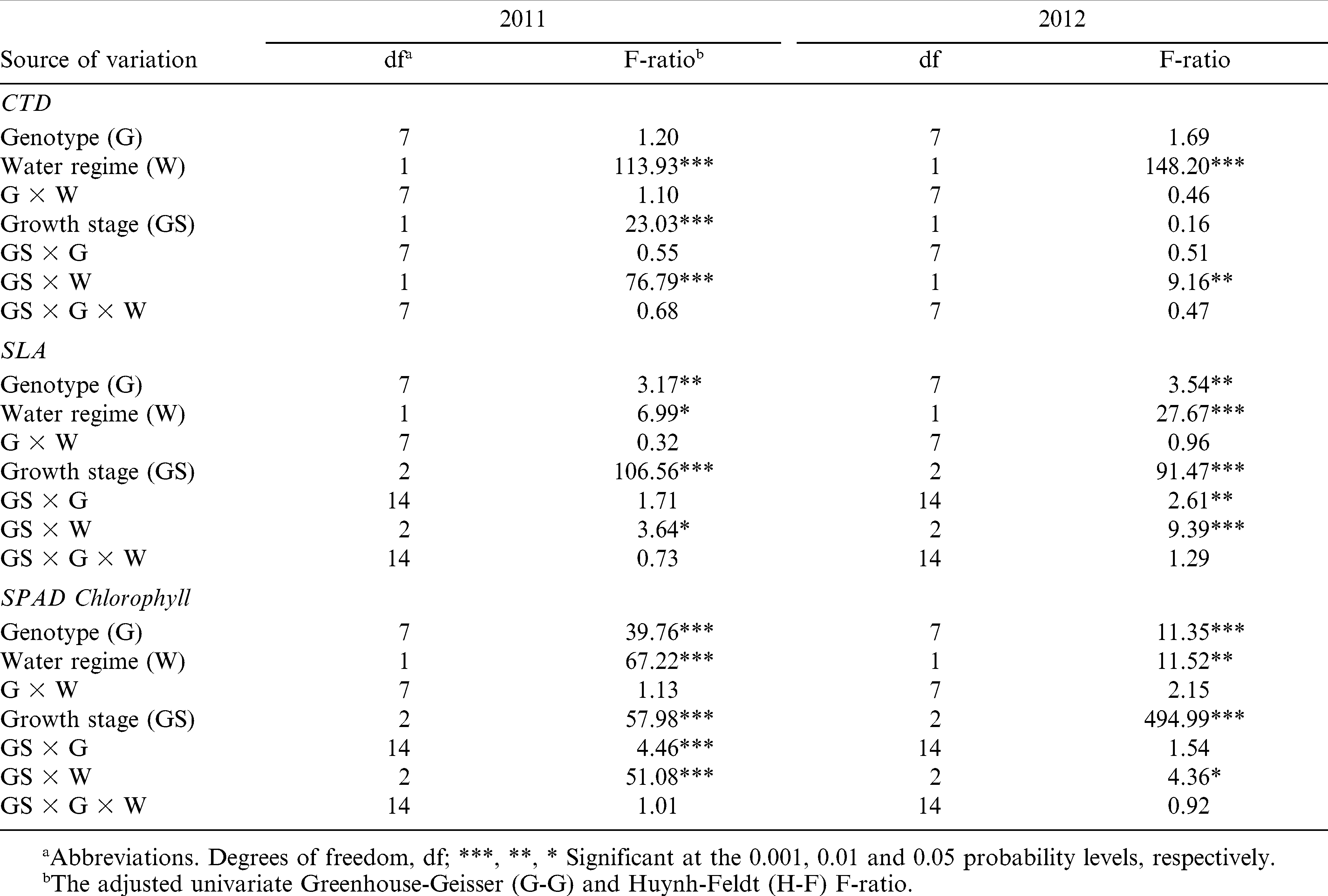
Univariate repeated measures analysis of canopy temperature depression (CTD), specific leaf area (SLA), and SPAD chlorophyll reading of eight peanut genotypes under two water regimes in 2011 and 2012.
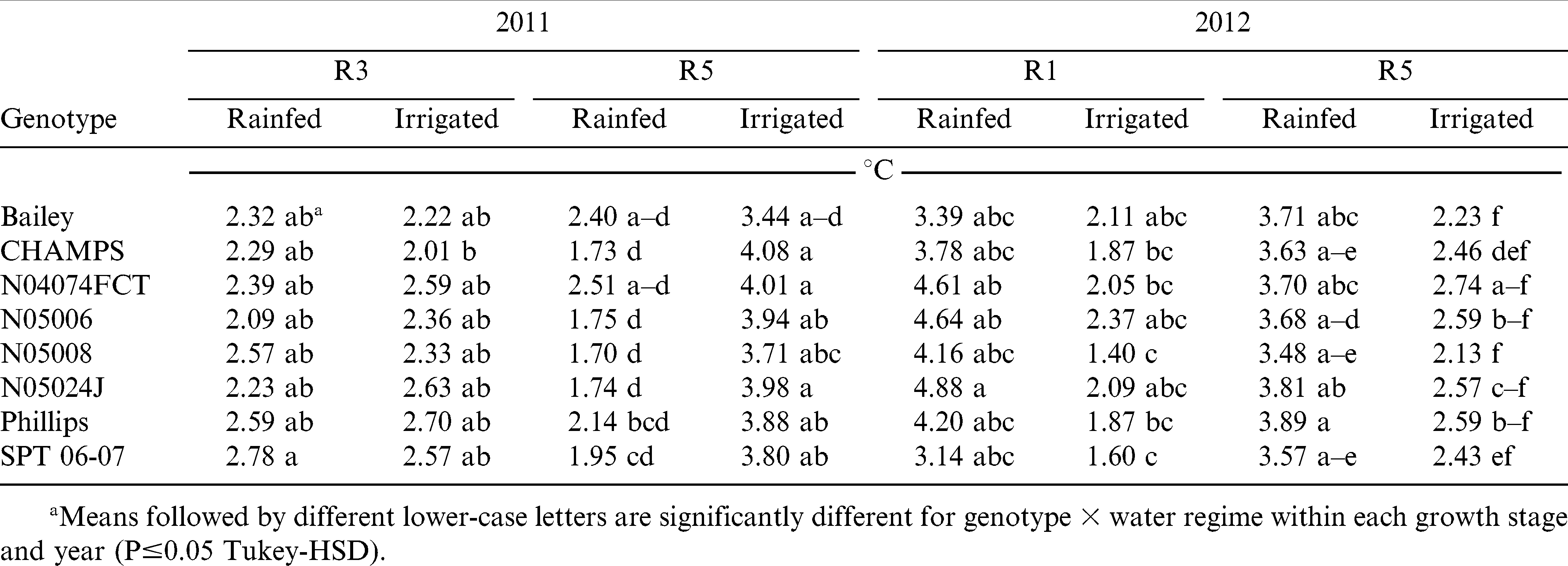
Canopy temperature depression (CTD) of eight virginia-type peanut genotypes under two water regimes (rainfed and irrigated) at two growth stages (R3, beginning pod; and R5, beginning seed) in 2011 and at three growth stages (R1, beginning flower and R5, beginning seed) in 2012 under field conditions.
Specific Leaf Area and SPAD Chlorophyll Reading
Genotype, water regime, and growth stage affected (p≤0.05) specific leaf area (SLA) and SPAD chlorophyll reading both years (Table 4). During both years, the growth stage by water regime interaction was significant (p≤0.05) for SLA and SPAD chlorophyll reading. In both years, the smallest SLA values were recorded at R3 (beginning pod) and the greenest leaves were at R5 (beginning seed) stages (Tables 6 and 7). Overall, leaves of rainfed plants had higher SLA (135 vs. 131 cm2 g−1) in 2011 and smaller SLA (133 vs. 144 cm2 g−1) in 2012 than the corresponding irrigated plants (Table 8). A similar pattern was observed for the SPAD chlorophyll readings. In 2011, average SPAD chlorophyll readings were 47.4 and 54.3 (relative units) for the rainfed and irrigated plants, respectively. In 2012, these numbers corresponded to 42.7 and 44.1 (relative units) for the rainfed and irrigated plants, respectively.
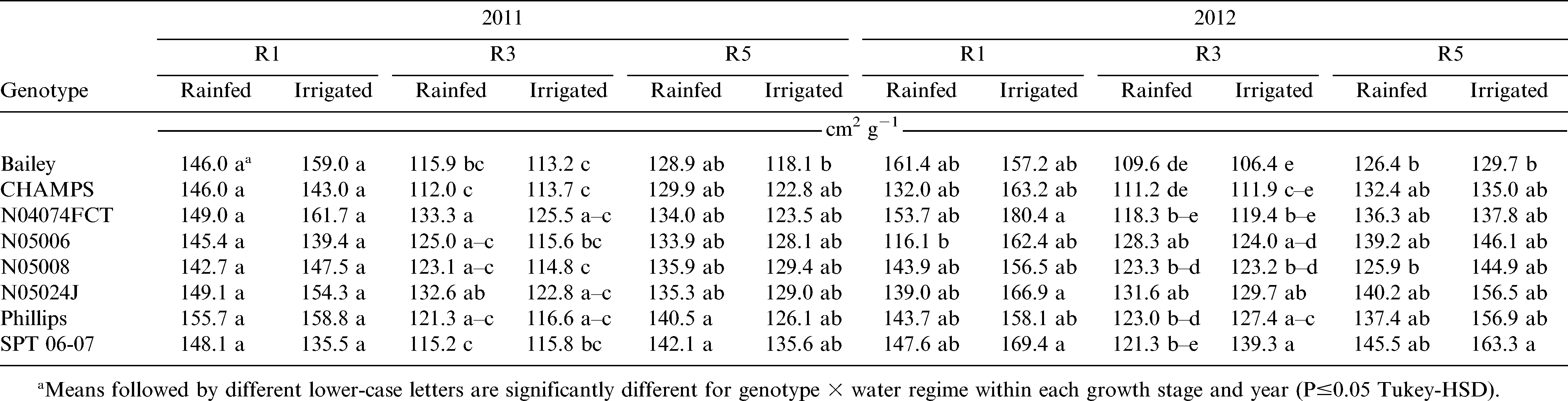
Specific leaf area (SLA) of eight virginia-type peanut genotypes under two water regimes (rainfed and irrigated) at three growth stages (R1, beginning flower; R3, beginning pod; and R5, beginning seed) in year 2011 and 2012 under field conditions.
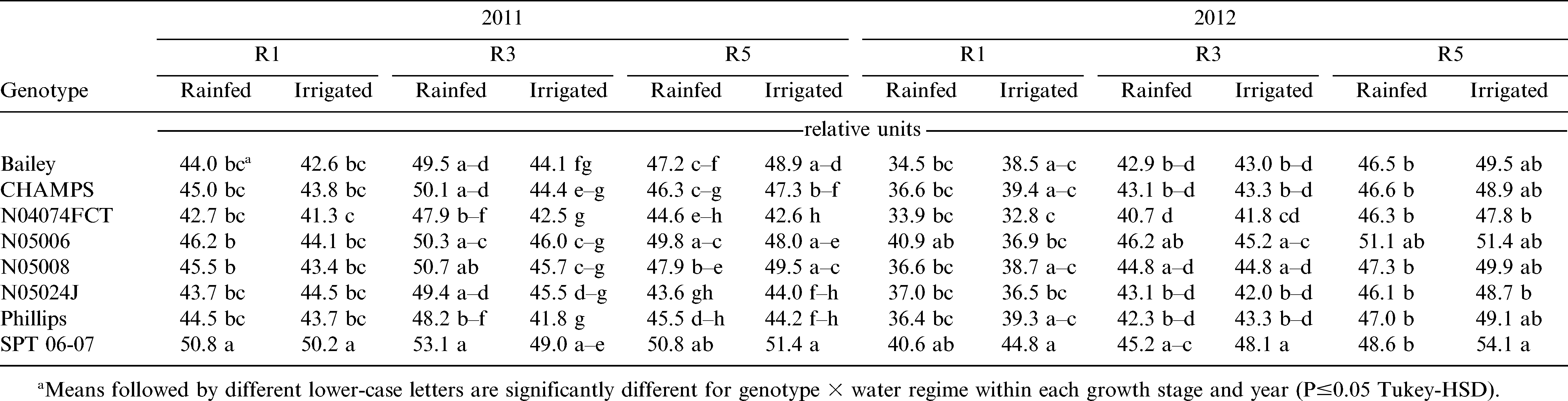
The SPAD chlorophyll reading of eight virginia-type peanut genotypes under two water regimes (rainfed and irrigated) at three growth stages (R1, beginning flower; and R5, beginning seed) in years 2011 and 2012 under field conditions.
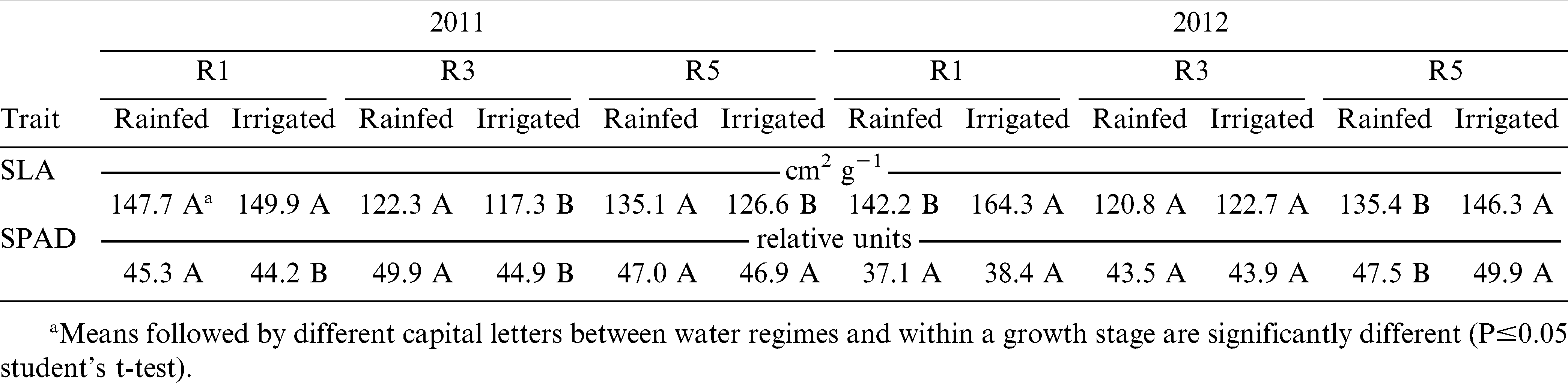
Mean specific leaf area (SLA) and SPAD chlorophyll reading of eight virginia-type peanut genotypes under two water regimes (rainfed and irrigated) at three growth stages (R1, beginning flower; R3, beginning pod; and R5, beginning seed) in year 2011 and 2012 under field conditions.
With a few exceptions, Bailey and CHAMPS had lower and Phillips higher SLA than other genotypes in both growing seasons (Table 6). For N04074FCT, SLA was relatively high at early stages and low at stage R5 in 2011 and 2012. N05006 and SPT 06-07 had lower SLA early than late in the season. Among all lines and based on the SPAD chlorophyll readings, SPT 06-07 was the greenest and N04074FCT the least green in both years and at all growth stages (Table 7).
Discussion
Pod Yield
Although soil moisture was adequate in 2011, i.e., total rainfall from May through October was 783 mm excluding the rainfall received from the tropical storm Irene, rainfed plots yielded on average 6% less than irrigated plots with a range of reduction due to water-deficit from less than 5% for Phillips and N05006 to approximately 7% for N05008, Bailey, CHAMPS, SPT 06-07, and N04074FCT (Table 3). The 2012 growing season was wetter than that of 2011 and quite hot based on historical data (Table 1). In addition to the 965 mm rainfall from May to October for all plots, irrigated plots received additional 32.3 mm irrigation from end of June to mid-July in 2012. Reduced radiation and excessive moisture could have contributed to lower yields in irrigated than in rainfed plots in 2012 as well as a 28% decrease in yield in 2012 compared to 2011 (Table 3).
Average pod yields varied significantly for different genotypes under rainfed conditions in both years. N05008, Phillips, and Bailey were the top yielding lines under rainfed conditions in 2011, while Phillips, N05024J, N05006, and N05008 were in 2012. In 2012, all genotypes showed increases in pod yield under rainfed conditions excepting only SPT 06-07, for which pod yield was significantly reduced (11.2%) under rainfed conditions. The lowest yielding lines in rainfed plots in both years were N04074FCT, CHAMPS, and SPT 06-07. Greenhouse studies of recovery from drought demonstrated that SPT 06-07 and N05006 recovered well from severe drought, while N04074FCT and Bailey showed a poor recovery from drought stress (Rosas-Anderson, 2012). Our field results are in an agreement with the greenhouse findings for N04074FCT and N05006, but not for Bailey and SPT 06-07. The discrepancies for Bailey and SPT 06-07 could be the result of their different yield potentials. Bailey has high yield potential that could have allowed this line to yield well under rainfed water regime even if the decrease from its irrigated yield was substantial, 7.9%. On the other hand, SPT 06-07 has inherently low yield potential, for which a 7.5% rainfed yield reduction could have made it the second lowest for yield after N04074FCT. Overall, Phillips, N05008, and N05006 were the best yielding lines under rainfed production in this study.
Canopy Temperature Depression
CTD has been widely used as a selection tool to screen germplasm for heat and drought tolerance in several crops (Amani et al., 1996; Balota et al., 2012a; Turner et al., 1986). The working principle of CTD is that under water stress, transpiration decreases due to stomata closure, but the tolerant genotypes tend to maintain higher transpiration rates, and therefore cooler canopies, than do the sensitive genotypes (Jackson, 1982; Turner et al., 1986). In the current study, no significant differences among genotypes were observed for CTD. Rainfed plants had on an average reduced CTD compared to irrigated plants (2.1 vs. 3.1 °C) in 2011, which was a dry year. In 2012, which was a wet year, irrigated plants were warmer (low CTD values) than rainfed plants (Table 5). This was probably due to excessive soil moisture in the irrigated plots that could have negatively affected the roots (Bradford and Hsiao, 1982; Jackson and Hall, 1987) resulting in reduced transpiration and therefore warmer plant canopies (reduced CTD). Canopy temperature depression was not correlated with pod yield in any water regime, growth stage, and year (Fig. 1). In 2012 under rainfed water regime, CTD measured early in the season (stage R1) showed a negative correlation with SLA (r = −0.44, p≤0.05). This suggests that reduction of SLA could be a physiological mechanism of peanut adaptation to rainfed production in sub-humid environments through better heat dissipation from thin leaves (higher SLA) when transpiration during the stress is low (Craufurd et al., 1999), but this needs to be confirmed by future observations. Absence of genotypic variation and correlation with pod yield suggests that CTD approach is not suited for genotypic comparisons for yield and water deficit tolerance in peanut grown in sub-humid environments.
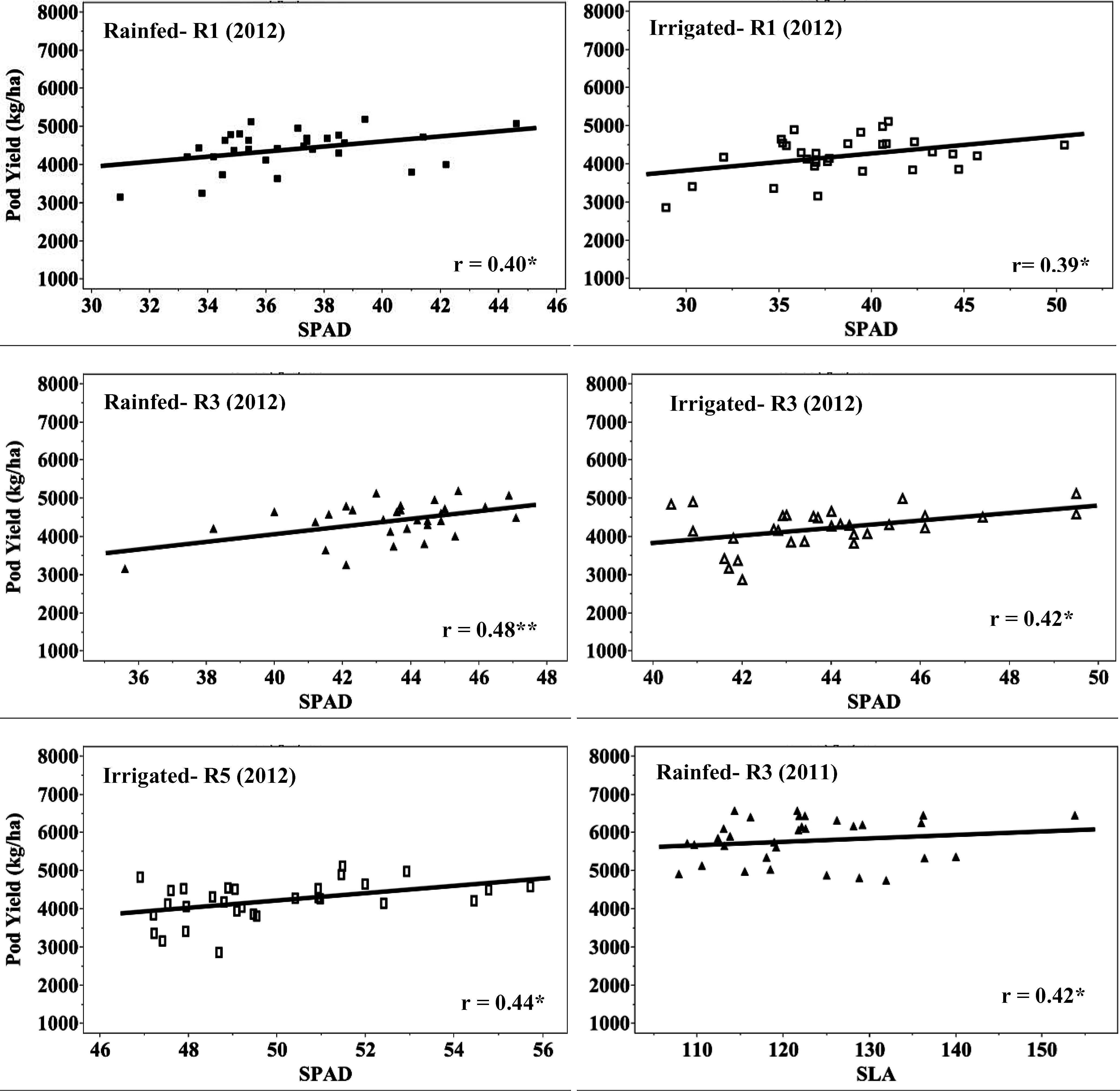
Correlation of pod yield, SPAD chlorophyll content, and SLA for eight virginia-type peanut genotypes at three growth stages (R1, beginning flower; R3, beginning pod; and R5, beginning seed), two water regimes (rainfed and irrigated) during field conditions in 2012.
Specific Leaf Area and SPAD Chlorophyll Reading
In both years, growth stage had a significant effect on the SLA and SPAD chlorophyll readings with the smallest SLA values being recorded at R3 (beginning pod), and the greenest leaves at R5 (beginning seed) (Tables 6 to 8). Overall, leaves from rainfed plants had higher SLA than irrigated plant leaves in 2011; but in 2012, leaves from irrigated plants had higher SLA than rainfed plant leaves (Table 8). Also, SPAD chlorophyll reading was higher in rainfed than irrigated plants in 2011, and higher in irrigated than in rainfed plants in 2012. Many studies report a decrease of the SLA in response to water deficit (Arunyanark et al., 2008; Upadhyaya, 2005), but others show an opposite trend (Wu et al., 2008). In our study, the observed increase of SLA can only be related to response to stress in general, if considering that irrigated plots in 2012 experienced excessive moisture and that plants under this water regime were more stressed than those in rainfed plots. There was no relationship between SLA and the SPAD chlorophyll reading in 2011, but a significant positive correlation was observed at stages R3 in rainfed (r = 0.37, p≤0.05) and irrigated (r = 0.50, p≤0.01) plots, and R5 in irrigated (r = 0.46, p≤0.01) plots in 2012 (Fig. 2). This can be explained by a considerable cloudiness in 2012, approximately 30% less incident radiation than in 2011, for which parallel increase of leaf area (high SLA) and chlorophyll content (better light capture) could have been necessary for increased photosynthesis and yield. There was no relationship between SLA and pod yield at any growth stage, water regime, and year. In 2012, SPAD chlorophyll readings at all three growth stages were significantly correlated to yield with coefficients of correlation ranging from 0.39 to 0.48 (Fig. 1). However, no relationship between yield and SPAD chlorophyll readings was detected in 2011.
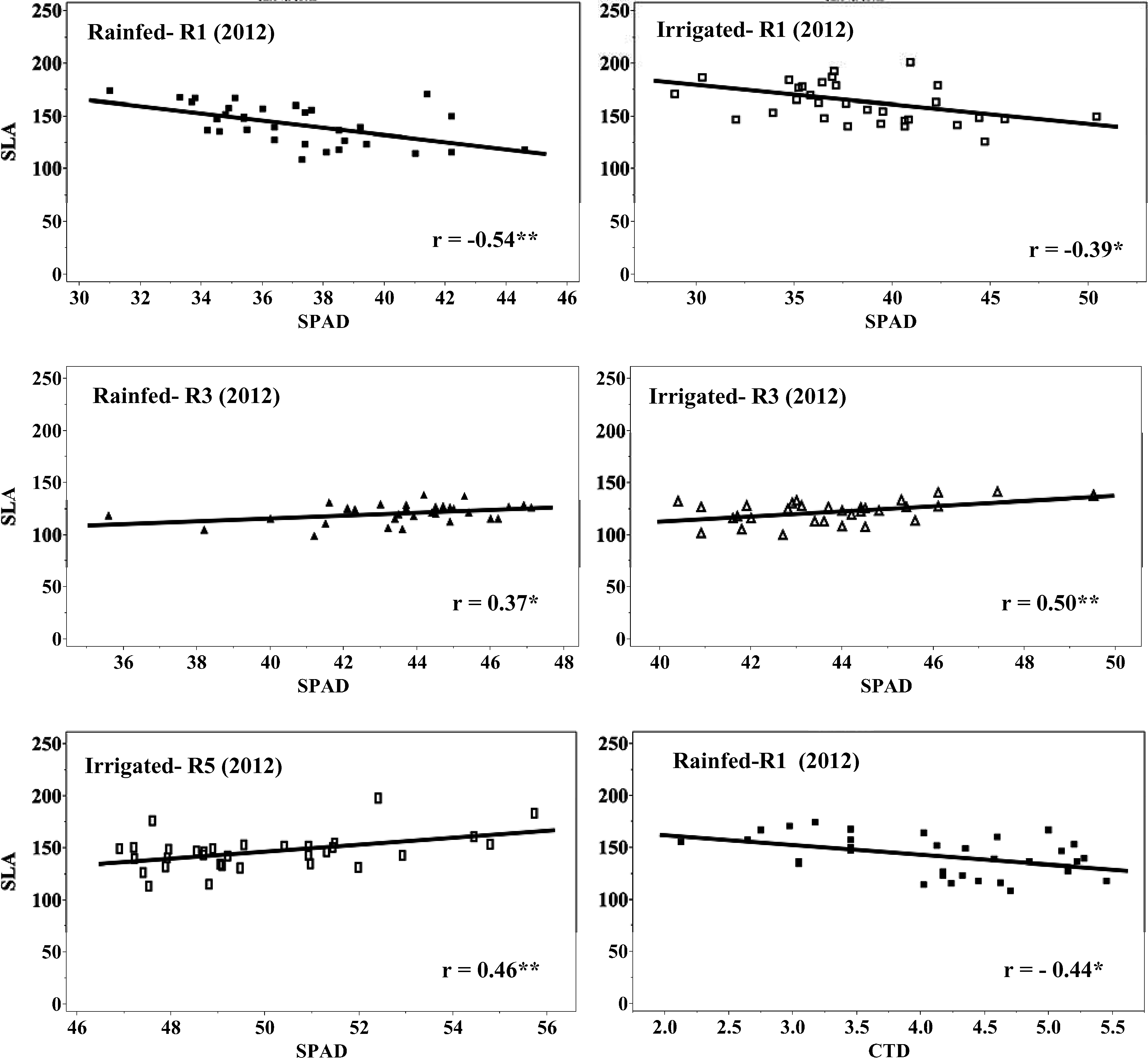
Correlation of CTD, SPAD chlorophyll content, and SLA for eight virginia-type peanut genotypes at three growth stages (R1, beginning flower; R3, beginning pod; and R5, beginning seed), two water regimes (rainfed and irrigated) during field conditions in 2012.
In summary and based on these results, none of the physiological characteristics measured in this study were consistently associated with yield and genotypic tolerance to water deficit in sub-humid climates where water stress level may be too low in some years for systematic field selection. Even though genotypic variability existed for SLA and SPAD chlorophyll reading, confirming our earlier findings ( Balota et al., 2012), their relationship with pod yield did not exist (CTD and SLA) or was limited to only one year (SPAD chlorophyll reading). This suggests that additional screening methods should be considered in sub-humid climates such as use of rain exclusion shelters or greenhouse coupled with field tests to increase the amount of drought stress in years and at growth stages when natural stress is absent.
Acknowledgements
This research project was funded by the National Peanut Board and Virginia Peanut Board, for which authors are very grateful. We are also grateful to Dr. Shyam Tallury for providing the seed of breeding line SPT 06-07.
Literature Cited
Amani I Fischer R.A and Reynolds M.P 1996 Canopy temperature depression association with yield of irrigated spring wheat cultivars in a hot climate J. Agron. Crop Sci. 176 : 119 – 129 .
Arunyanark A Jogloy S Akkasaeng C Vorasoot N Kesmala T Nageswara Rao R.C Wright G.C and Patanothai A 2008 Chlorophyll stability is an indicator of drought tolerance in peanut J. Agron. Crop Sci. 194 : 113 – 125 .
Ayeneh A van Ginkel M Reynolds M.P and Ammar K 2002 Comparison of leaf, spike, peduncle and canopy temperature depression in wheat under heat stress Field Crops Res. 79 : 173 – 184 .
Balota M Payne W.A Evett S.R and Peters T.R 2008 Morphological and physiological traits associated with canopy temperature depression in three closely related wheat lines Crop Sci. 48 : 1897 – 1910 .
Balota M Isleib T.G and Tallury S 2012a Variability for drought related traits of virginia-type peanut cultivars and advanced breeding lines Crop Sci. 52 : 2702 – 2713 .
Balota M McGrath S Isleib T.G and Tallury S 2012b Transpiration response to vapor pressure deficit in field grown peanut Peanut Sci. 39 : 53 – 61 .
Bell M.J Roy R.C Tollenaar M and Michaels T.E 1994 Importance of variation in chilling tolerance for peanut genotypic adaptation to cool, short-season environments Crop Sci. 34 : 1030 – 1039 .
Blum A Mayer J and Golan G 1988 The effect of grain number per ear (sink size) on source activity and its water-relations in wheat J. Exp. Bot. 39 : 106 – 114 .
Boote K.J 1982 Growth stages of peanut (Arachis hypogaea L.) Peanut Sci. 9 : 35 – 40 .
Bradford K.J and Hsiao T.C 1982 Stomatal behavior and water relations of waterlogged tomato plants Plant Physiol. 70 : 1508 – 1513 .
Chapman S.C and Barreto H.J 1997 Using a chlorophyll meter to estimate specific leaf nitrogen of tropical maize during vegetative growth Agron. J. 89 : 557 – 562 .
Collaku A and Harrison S.A 2002 Losses in wheat due to waterlogging Crop Sci. 42 : 444 – 450 .
Craufurd P.Q Wheeler T.R Ellis R.H Summerfield R.J and Williams J.H 1999 Effect of temperature and water deficit on water-use efficiency, carbon isotope discrimination, and specific leaf area in peanut Crop Sci. 39 : 136 – 142 .
FAOSTAT 2013 , Statistical Data Base Food and Agricultural Organizations of the United Nations .
Isleib T.G
Milla-Lewis S.R
Pattee H.E
Copeland S.C
Zuleta M.C
Shew B.B
Hollowell J.E
Sanders T.H
Dean L.O
Hendrix K.W
Balota M
and
Chapin J.W
2011
Registration of ‘Bailey’ peanut
J. Plant Reg.
5
:
27
–
39
(doi:10.3198/jpr2009.12.0742crc)
Isleib T.G Rice P.W Mozingo R.W Copeland S.C Graeber J.B Pattee H.E Sanders T.H Mozingo R.W and Coker D.L 2006 Registration of ‘Phillips’ peanut Crop Sci. 46 : 2308 – 2309 .
Jackson M.B and Hall K.C 1987 Early stomatal closure in waterlogged pea plants is mediated by abscisic acid in the absence of foliar water deficits Plant Cell Environ. 10 : 121 – 130 .
Jackson R.D 1982 Canopy temperature and crop water stress Adv. Irrig. 1 : 43 – 85 .
Mozingo R.W Coffelt T.A Phipps P.M and Coker D.L 2006 Registration of ‘CHAMPS’ peanut Crop Sci. 46 : 2711 – 2712 .
Nageswara Rao R.C Talwar H.S and Wright G.C 2001 Rapid assessment of specific leaf area and leaf nitrogen in peanut (Arachis hypogaea L.) using a chlorophyll meter J. Agron. Crop Sci. 186 : 175 – 182 .
O'Toole J.C Turner N.C Namuco O.P Dingkuhn M and Gomez K.A 1984 Comparison of some crop water stress measurement methods Crop Sci. 24 : 1121 – 1128 .
Reynolds M Manes Y Izanloo A and Langridge P 2009 Phenotyping approaches for physiological breeding and gene discovery in wheat Ann.Appl.Biol. 155 : 309 – 320 .
Rorie R.L Purcell L.C Mozaffari M Karcher D.E King C.A Marsh M.C and Longer D.E 2011 Association of “greenness” in corn with yield and leaf nitrogen concentration Agron. J. 103 : 529 – 535 .
Rosas-Anderson P.J 2013 Peanut genetic variation for recovery from severe soil drying and low epidermal conductance as a drought tolerance trait. M.S. thesis North Carolina State University , Raleigh .
SAS Institute Inc
2012
,
SAS Institute Cary, NC, USA.
Sheshshayee M.S Bindumadhava H Rachaputi N.R Prasad T.G Udayakumar M Wright G.C and Nigam S.N 2006 Leaf chlorophyll concentration relates to transpiration efficiency in peanut Ann. Appl. Biol. 148 : 7 – 15 .
Songsri P Jogloy S Holbrook C.C Kesmala T Vorasoot N Akkasaeng C and Patanothai A 2009 Association of root, specific leaf area and SPAD chlorophyll meter reading to water use efficiency of peanut under different available soil water Agr. Water Manage. 96 : 790 – 798.
Turner N.C O'Toole J.C Cruz R.T Namuco O.S and Ahmad S 1986 Responses of seven diverse rice cultivars to water deficits I. Stress development, canopy temperature, leaf rolling and growth Field Crops Res. 13 : 257 – 271 .
Upadhyaya H.D 2005 Variability for drought resistance related traits in the mini core collection of peanut Crop Sci. 45 : 1432 – 1440 .
Wu F.Z Bao W.K Li F.L and Wu N 2008 Effects of water stress and nitrogen supply on leaf gas exchange and fluorescence parameters of Sophora davidii seedlings Photosynthetica 46 : 40 – 48 .
Notes
- Graduate Research Assistant, Department of Plant Pathology, Kansas State University, Manhattan, KS 66506
- Assistant Professor, Tidewater Agricultural Research and Extension Center, Virginia Polytechnic Institute and State University, Suffolk, VA 23437
- Professor, Department of Crop Science, North Carolina State University, Raleigh, NC 27695
- Assistant Professor, Department of Plant Pathology, Physiology and Weed Science, Virginia Polytechnic Institute and State University, Blacksburg, VA 24061
- Professor, Horticulture Department, Virginia Polytechnic Institute and State University, Blacksburg, VA 24061 * Corresponding author email: singhdj2@ksu.edu
Author Affiliations