

Introduction
Concerns regarding glyphosate-resistant weeds has led to an interest in developing alternative herbicide-tolerant crops. Dicamba-tolerance is being developed in several broadleaf crops including soybean [Glycine max (L.) Merr.] and cotton (Gossypium hirsutum L.) (Behrens et al. 2007; Subramanian et al. 1997). Currently, dicamba is registered for postemergence broadleaf weed control use in various grass crops such as field corn (Zea mays L.), sorghum [Sorghum bicolor (L.) Moench], and wheat (Triticum aestivum L.) (Anonymous, 2011).
Dicamba's reputation for off -target movement due to drift and volatility has been well documented (Al-Khatib and Peterson, 1999; Behrens and Lueschen, 1979). In the southeast, peanut is grown in close proximity to both soybean and cotton. Thus, the adoption of dicamba-tolerance in these crops increases the probability of drift, volatilization, and tank contamination problems that could negatively influence peanut development and yield.
Peanut response to dicamba has not been well documented. Dicamba applied at approximately 2 g/ha had no effect on peanut yield in one field trial (Prostko et al. 2009). However, other studies on the control of volunteer peanut indicated that peanut is not tolerant of dicamba (York et al. 1994). In a related forage crop, rhizome peanut (Arachis glabrata Benth) yields were significantly reduced by a foliar application of dicamba + 2,4-D (Ferrell et al. 2006). Other systematic studies on the influence of dicamba rate and timing on peanut have not been published in the literature. Therefore, the objective of this research was to quantify the effects of various rates of dicamba, applied at 30, 60 or 90 days after planting (DAP), on peanut yield.
Materials and Methods
Field trials were conducted at eight locations across the United States peanut belt during 2008. A complete description of these locations is presented in Table 1. Production and pest management practices were followed according to local Cooperative Extension recommendations.
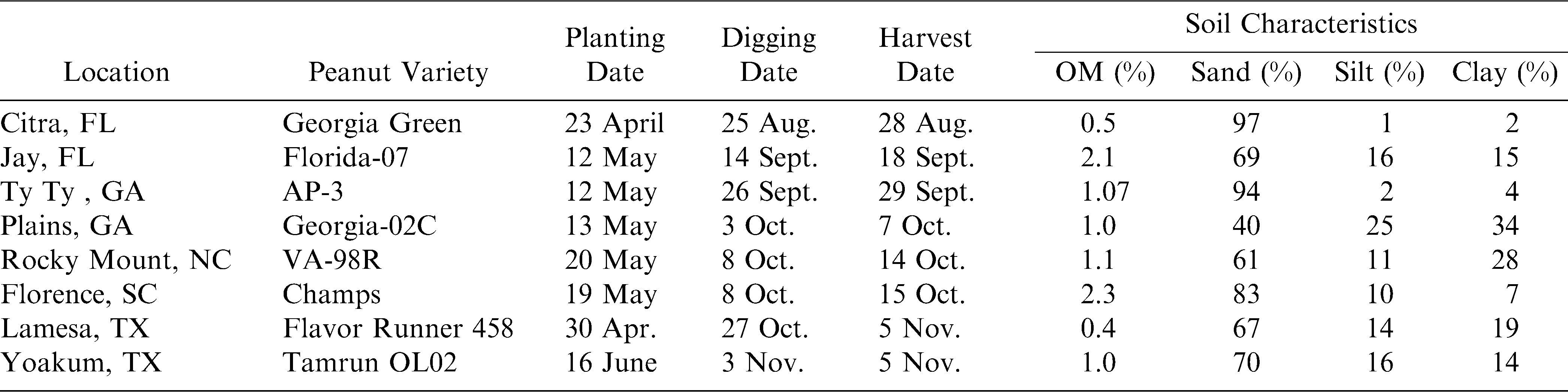
Locations, varieties, planting dates, harvest dates, and soil types for the peanut/dicamba tolerance studies in 2008.
All trials were conducted in a randomized complete block design with a three (application timing) by six (dicamba rate) factorial arrangement of treatments. Dicamba timings were 30, 60, and 90 DAP and dicamba rates were 0, 40, 70, 140, 280, 560 g ai/ha. The typical use rate of dicamba in grass crops is 280 g ai/ha. All treatments were replicated four times. Dicamba was applied using a CO2-pressurized sprayer calibrated to deliver 94–140 L/ha. All plot areas were maintained weed-free throughout the season using a combination of herbicides (pendimethalin, diclosulam, flumioxazin, imazapic, 2,4-DB) and hand-weeding. Peanut yield data were obtained by mechanical harvesting at maturity.
The data were analyzed as a split-plot. Location was the main plot with herbicide rate and days after planting randomized within each location.
Dependent variables were transformed to percent of control then subtracted from 100 to represent percent loss. PROC GLM (SAS, 2008) was used to analyze the full model. When there were significant interactions between location and rate or location and timing, rate and timing were analyzed within each location. Regression models were developed by regressing percent yield loss over herbicide rate. Linear models were fit using SigmaPlot1 and the results are summarized in Figures 1– 4.
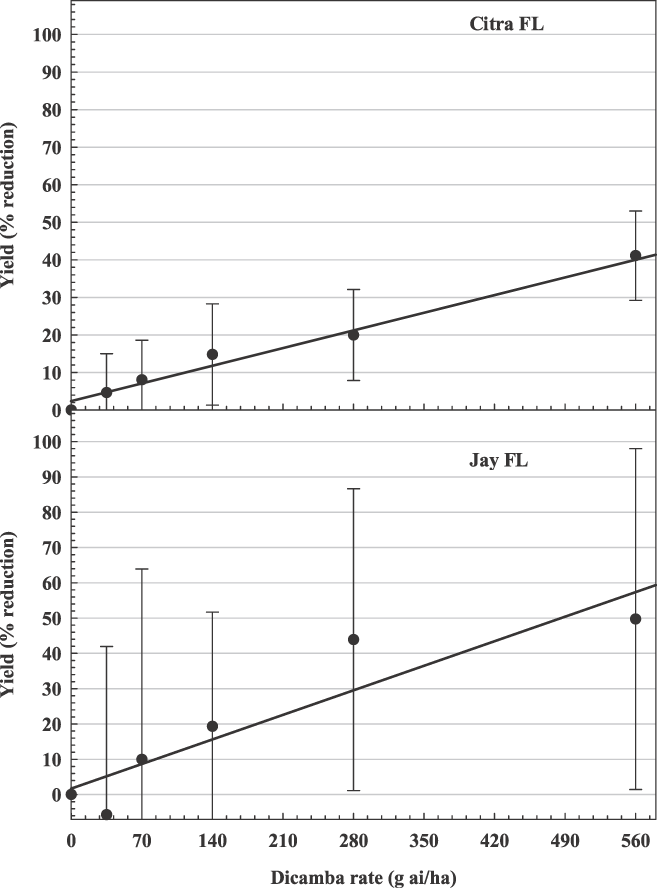
The influence of dicamba rate on peanut yield loss averaged over 3 application timings (30, 60, and 90 DAP) in Florida. Citra, y = 0.07x + 2.37, R2 = 0.61; Jay, y = 0.10x + 1.72, R2 = 0.18.
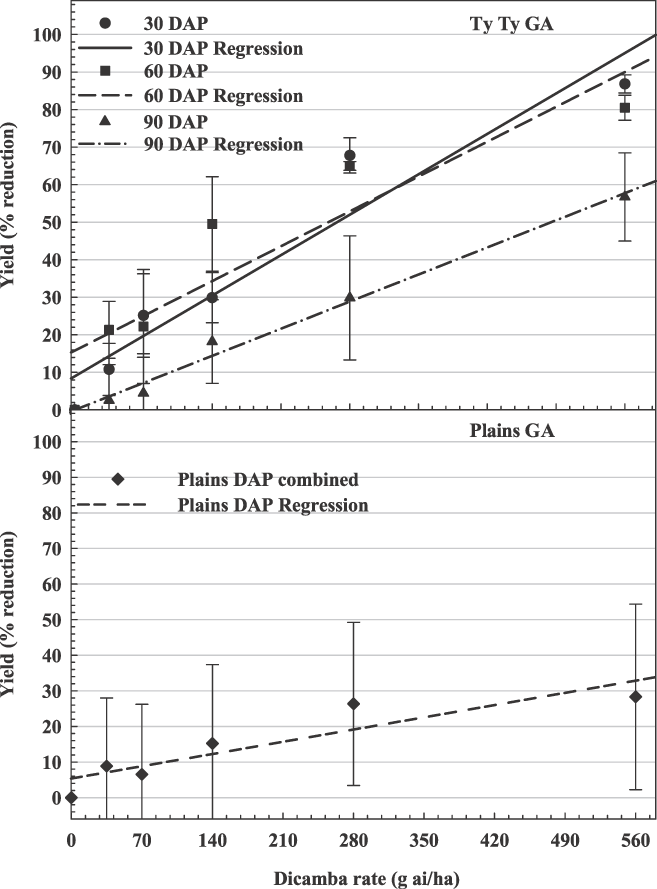
The influence of dicamba rate on peanut yield loss in Georgia. Ty Ty, 30 DAP, y = 0.15x + 8.93, R2 = 0.89; 60 DAP, y = 0.13x + 15.78, R2 = 0.78; 90 DAP, y = 0.10x − 0.03, R2 = 0.80; Plains (averaged over 3 application timings), y = 0.05x + 5.37, R2 = 0.18.
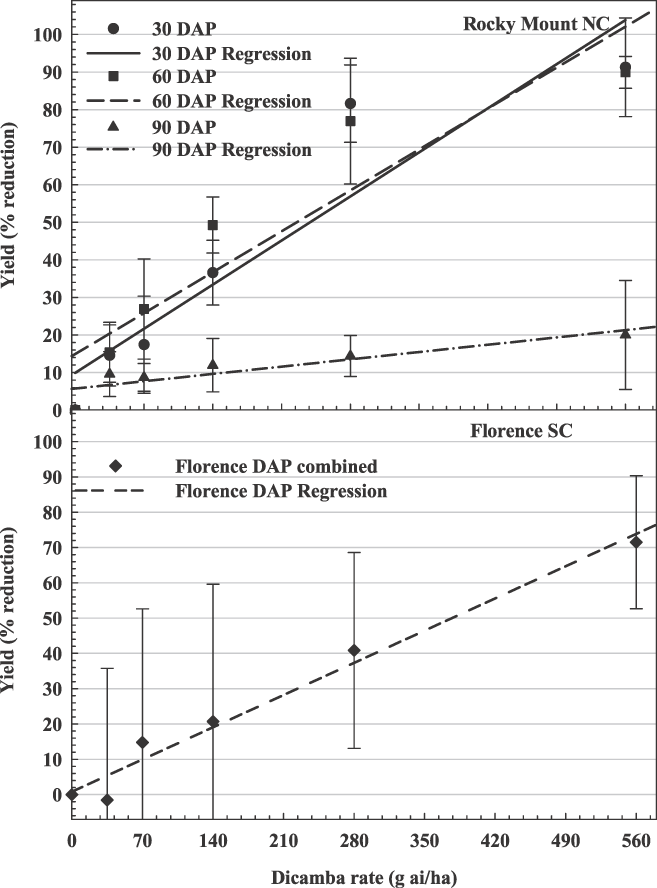
The influence of dicamba rate on peanut yield loss in North and South Carolina. Rocky Mount, 30 DAP, y = 0.17x + 9.91, R2 = 0.82; 60 DAP, 0.16x + 14.92, R2 = 0.80; 90 DAP, 0.03x + 5.74, R2 = 0.36; Florence (averaged over 3 application timings), y = 0.13x + 0.76, R2 = 0.43.
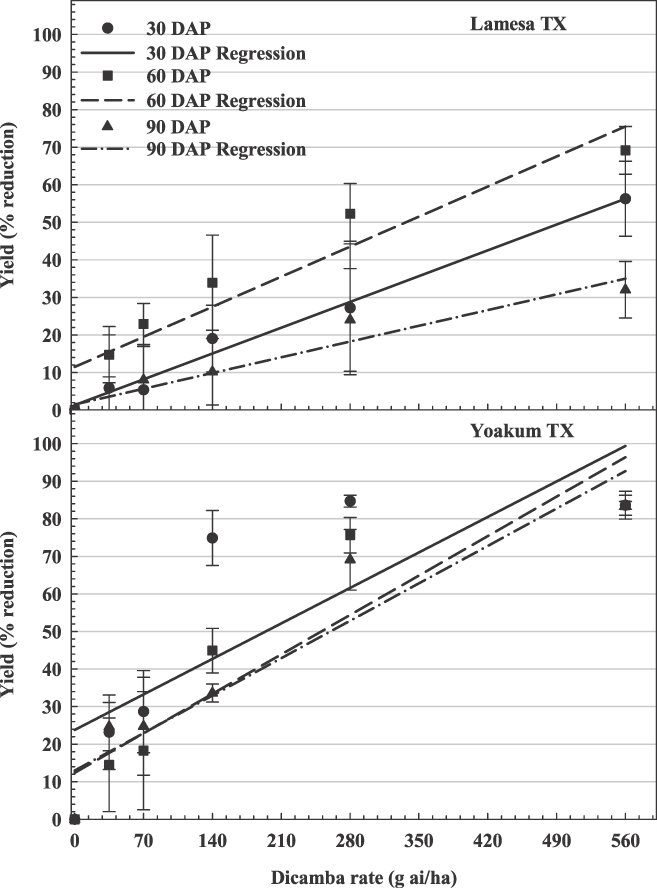
The influence of dicamba rate on peanut yield loss in Texas. Lamesa, 30 DAP, y = 0.10x + 1.33, R2 = 0.75; 60 DAP, y = 0.11x + 11.50, R2 = 0.84; 90 DAP, y = 0.06x + 1.50, R2 = 0.67; Yoakum, 30 DAP, y = 0.14x + 23.79, R2 = 0.61; 60 DAP, 0.15x + 12.36, R2 = 0.79; 90 DAP, 0.14x + 12.97, R2 = 0.85.
Results and Discussion
Significant interactions between location and dicamba timing prevented pooling of data (P = 0.0001). Therefore, data is presented by individual location. ANOVA tables by locations are presented in Table 2.
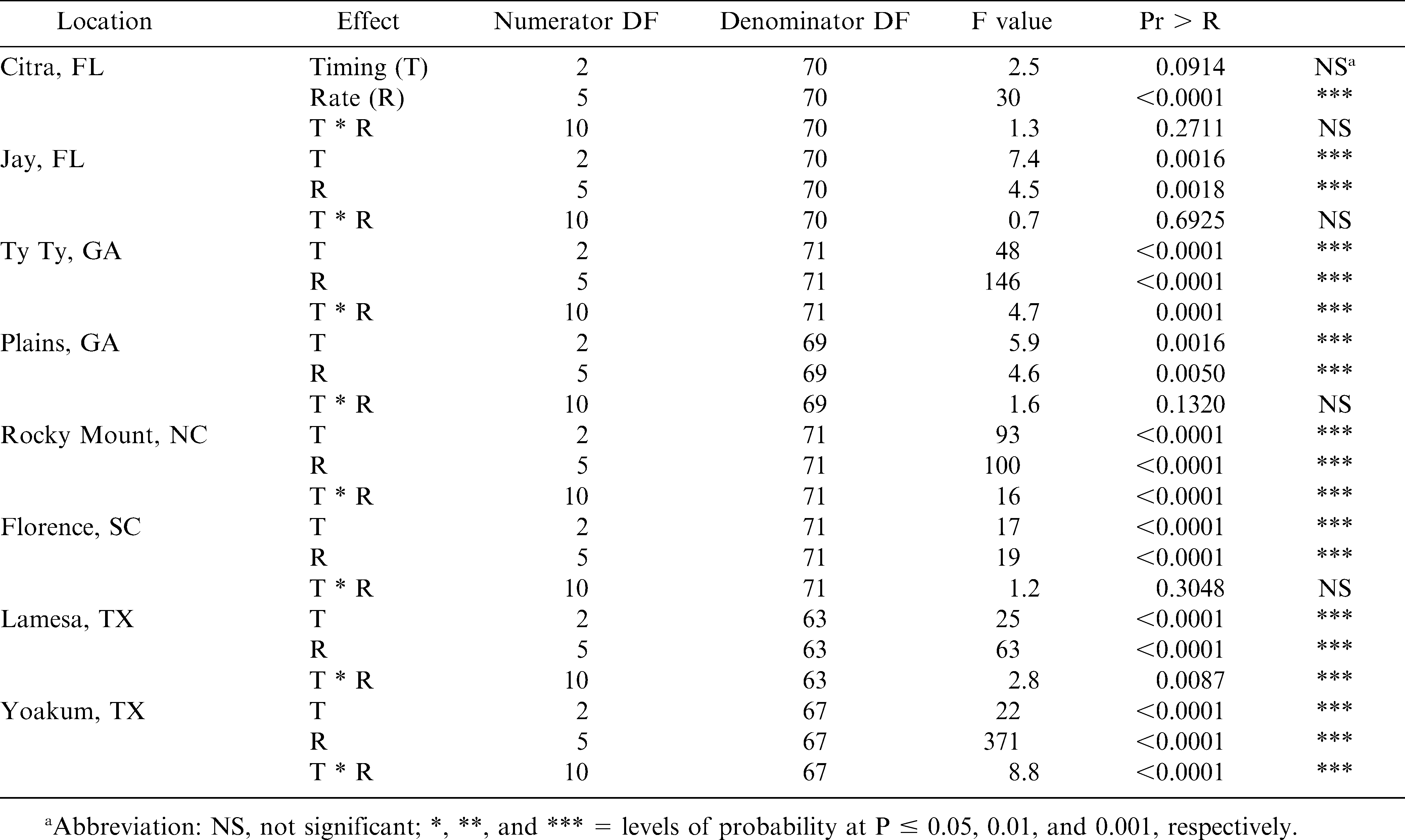
Analysis of variance for dicamba effects on peanut pod yield loss in Florida, Georgia, North Carolina, South Carolina, and Texas.
At Citra, Jay, Plains, and Florence, there were no interactions between dicamba rate and timing. At these locations, there was a weak linear relationship (R2 range from 0.18–0.61) between dicamba rate and peanut yield loss when pooled over application timing (Figures 1, 2, and 3). Generally, as dicamba rate increased, peanut yield decreased. Estimated peanut yield losses from the lowest rate of dicamba (40 g/ha) were 5%, 6%, 7%, 6% at Citra, Jay, Plains, and Florence, respectively. Estimated peanut yield losses from the highest rate of dicamba (560 g/ha) were 42%, 58%, 33%, 74%, at Citra, Jay, Plains, and Florence, respectively.
At the Citra location, dicamba timing had no effect on peanut yield loss (Table 3). At Jay, applications of dicamba at 30 DAP caused greater yield losses than when applied at 60 or 90 DAP. In Plains, dicamba applied at 60 and 90 DAP resulted in greater yield losses than when applied at 30 DAP. In Florence, dicamba applied at 30 and 60 DAP, resulted in greater yield losses than when applied at 90 DAP.

Peanut yield loss as influenced by dicamba timing at four locations (Citra, Jay, Plains, Florence).ab
At Ty Ty, Rocky Mount, Lamesa, and Yoakum, there were significant interactions between dicamba rate and time of application (Figures 2, 3, and 4). At these locations, there was a stronger linear relationship between dicamba rate and yield loss (R2 range from 0.36–0.89). Generally, peanut yield losses were greater when dicamba was applied at 30 and 60 DAP when compared to 90 DAP. Peanut may be more tolerant of dicamba at 90 DAP for several reasons. First, significant vine density at this time may impede spray coverage. Second, approximate stages of peanut growth were R1 (beginning bloom), R3 (beginning pod) and R6 (full seed) at 30, 60, and 90 DAP, respectively. In soybean, dicamba caused greater yield reductions when exposure occurred at bloom (Auch and Arnold, 1978; Wax et al., 1969; Weidenhamer et al., 1989). Similar results have also been observed in cotton (Hamilton and Arle 1979). Estimated yield losses from the lowest rate of dicamba (40 g/ha) were 2% to 29%, 16% to 21%, and 4% to 19% for 30, 60, and 90 DAP, respectively. Estimated yield losses from the highest rate of dicamba (560 g/ha) were 57% to 100%, 73% to 100%, and 23% to 91% for 30, 60, and 90 DAP, respectively.
The original intent of this research was to develop a single model that could be used across the U.S. peanut belt to accurately estimate the potential negative yield effects of off-target movement or sprayer contamination of dicamba. Unfortunately, the variability in peanut response between locations was greater than anticipated. It is reasonable to assume that the differences in peanut yield loss caused by dicamba rate and timing between locations was related to variety, soil type, and environmental conditions. Regardless of the specific results at these locations, it is evident that peanut is not tolerant of dicamba. Additionally, these results further justify the continued need for state specific weed science research.
Summary and Conclusions
Significant peanut yield losses from dicamba were observed at rates as low as 40 g/ha (0.14× normal use rate). Yield losses from this rate ranged between 2% to 29%. Generally, peanut plants were more sensitive to dicamba at 30 and 60 DAP. If the use of dicamba-tolerant cotton and soybean becomes widely adopted in the future, peanut growers must avoid dicamba drift and sprayer contamination, particularly during the early reproductive stages of growth.
Sources of Materials
1SigmaPlot, Version 11, Systat Software Inc., 1735 Technology Drive, Suite 430, San Jose, CA 95110.
Acknowledgements
This research was partially supported through a grant provided by the Georgia Peanut Commission, North Carolina Peanut Grower's Association, and the Texas Peanut Producer's Board. The technical support of Charlie Hilton, Jesse Parker, Scott Clewis, Dewayne Johnson, and Lyndell Gilbert was greatly appreciated.
Literature Cited
Al-Khatib K. and Peterson D. 1999 Soybean (Glycine max) response to simulated drift from selected sulfonylurea herbicides, dicamba, glyphosate, and glufosinate. Weed Technol 13 : 264 – 270 .
Anonymous 2011 Clarity ® product label BASF Corporation Research Triangle Park, NC .
Auch D. E. and Arnold W. E. 1978 Dicamba use and injury on soybeans (Glycine max) in South Dakota. Weed Sci 26 : 471 – 475 .
Behrens R. and Lueschen W. E. 1979 Dicamba volatility. Weed Sci 27 : 486 – 493 .
Behrens M. R. , Mutlu N. , Chakraborty S. , Dumitru R. , Zhi-Jang W. , LaVallee B. J. , Herman P. L. , Clemente T. E. , and Weeks D. P. 2007 Dicamba resistance: Enlarging and preserving biotechnology-based weed management strategies. Science 316 : 1185 – 1188 .
Ferrell J. A. , Sellers B. A. , Mudge C. R. , and Smith C. A. 2006 Evaluation of postemergent herbicides on rhizoma peanut injury and yield. Online. Forage and Grazinglands doi:10.1094/FG-2006-0308-01-RS.
Hamilton K. C. and Arle H. F. 1979 Response of cotton (Gossypium hirsutum) to dicamba. Weed Sci 27 : 604 – 607 .
Prostko E. P. , Flanders J. T. , and Brown S. N. 2009 Extension response to contaminated 2,4-DB/peanut problems in 2007. Weed Sci. Soc. Amer. Abstr 49 : 358 .
SAS Institute Inc 2008 SAS OnlineDoc(r) 9.2 Cary, NC SAS Institute Inc .
Subramanian M. V. , Tuckey J. , Patel B. , and Jensen P. J. 1997 Engineering dicamba selectivity in crops: a search for appropriate degradative enzyme(s). J. of Indus. Micro. & Biotech 9 : 344 – 349 .
Wax L. M. , Knuth L. A. , and Slife F. W. 1969 Response of soybean to 2,4-D, dicamba, and picloram. Weed Sci 17 : 388 – 393 .
Weidenhamer J. D. , Triplett G. B. , and Sobotka F. E. 1989 Dicamba injury to soybean. Agron. J 81 : 637 – 643 .
York A. C. , Jordan D. L. , and Wilcut J. W. 1994 Peanut control in rotational crops. Peanut Sci 21 : 40 – 43 .
Notes
- Prof., Dept. Crop & Soil Sci., The Univ. of Georgia, Tifton, GA 31793. [^]
- Assoc. Prof., Dept. Crop & Soil Sci., The Univ. of Georgia, Tifton, GA 31793. [^]
- Asst. Prof., Dept. Entom., Soils and Plant Sci., Clemson Univ., Blackville, SC 29817. [^]
- Assoc. Prof., Agron. Dept., Univ. of Florida, Gainesville, FL 32611. [^]
- Prof., Dept. Plant & Soil Sci., Texas Tech Univ., Lubbock, TX 79409. [^]
- Prof., Crop Sci. Dept., North Carolina State Univ., Raleigh, NC 27695. [^]
- Senior. Res. Scientist, Texas AgriLife Research, Beeville, TX 77802. [^]
- Prof., Agron. Dept., Univ. of Florida, Jay, FL 32565. [^]
- Ag. Res. Statistician, The Univ. of Georgia, Griffin, GA 30223. [^] *Corresponding author (email: eprostko@uga.edu)
Author Affiliations