

Introduction
Sclerotinia blight of peanut (Arachis hypogaea L.), caused by the soil-borne fungal pathogen Sclerotinia minor Jagger, was first reported in the southeastern U.S. in Virginia in 1971 (Porter and Beute, 1974) and has been reported in other peanut producing states including North Carolina, Oklahoma (Wadsworth, 1979) and Texas (Woodard and Simpson, 1993). Losses in fields affected by Sclerotinia blight can approach 50% (Porter and Melouk, 1997). Ideal management of the disease is to exclude the pathogen from new fields. Once a field is infested, effective chemical management is very expensive and a broad host range limits the benefits of long rotations. A rotation of peanut for 1 year followed by 3 years of cotton and then peanut again may result in considerable disease caused by S. minor (T. Wheeler, personal observations).
S. minor infects peanut tissue primarily through mycelia produced by germinating sclerotia during periods of cool, moist conditions (Porter and Melouk, 1997). S. minor can infect all parts of the peanut plant, however stems are primarily affected. Mycelia of S. minor often appear as a white cottony mass on the surface of infected tissue. As the disease progresses, water soaked stem lesions will become bleached in appearance and eventually become necrotic. Small (0.5 to 2 mm), highly melanized, irregular shaped sclerotia are formed abundantly on and in dead tissue. Sclerotia can be sloughed from tissue or remain attached to old plant parts after harvest and can spread from field to field through peanut seed, windblown soil and plant tissue, farm machinery, and workers (Akem and Melouk, 1990; Porter and Melouk, 1997). S. minor sclerotia are capable of long-term survival in soil, particularly under dry conditions (Wu et al., 2008) such as those prevalent in the semiarid environment of west Texas.
Sodium hypochlorite (NaOCl) has been used by producers to clean equipment and clothing infested with sclerotia of S. minor. However, little is known about optimal concentration and exposure time necessary to kill sclerotia of S. minor. Linderman and Gilbert (1972) found that sclerotia of Sclerotium rolfsii Sacc. were still viable after 30 min in 0.5% NaOCl. The objective of this project was to determine the exposure time and NaOCl concentration required to kill sclerotia of S. minor.
Materials and Methods
An isolate of S. minor was obtained by culturing sclerotia obtained from diseased peanut stems from Gaines Co., TX and was maintained on potato-dextrose agar (PDA) at room temperature (20-to-23 C). Mature sclerotia were obtained directly from 3- to 4-week-old PDA cultures.
Effect of Sodium Hypochlorite on Bare Sclerotia
Sclerotia were placed in 200 ml beakers containing 150 ml of 0, 0.6, 3.0, or 6.0% NaOCl (Clorox® bleach, Oakland, CA) solution. Sclerotia were stirred in solution for 2, 5, or 10 min. Approximately 1 min after stirring; half of the total number of sclerotia were placed on a sieve with a 700 μm opening and rinsed for 10 sec with a wash bottle containing sterile deionized water. Sclerotia were then air dried under a laminar flow hood for 2 min. PDA plates (100 × 15 mm) were divided into 4 quadrants and a treated sclerotium was placed in the center of each quadrant for a total of 16 sclerotia per treatment. The growth of mycelia (maximum diameter 5 cm) from each sclerotium was measured after 7 days. Plates with no germination were observed for several weeks to ensure that the sclerotia were dead. Plates were arranged on a laboratory bench at room temperature in a randomized complete block design with 4 replications. Data recorded were binomial, where any growth of mycelia from a single sclerotia was recorded as 1, while no growth was recorded as 0. The experiment was repeated once.
Effect of Sodium Hypochlorite on Sclerotia in Soil
Sixteen sclerotia were added to 50 ml of a 5:1 soil:NaOCl mixture (v/v). Soil types were a sandy clay loam (52% sand, 26% silt, 22% clay, 0.7% OM) from Lubbock, Co. and a sandy loam (72% sand, 10% silt, 18% clay, 0.3% OM) from Gaines Co. Sclerotia were added to the mixture and stirred by hand for 30 s then allowed to settle for 20, 40, 60, or 80 min. Treated sclerotia were retrieved on a sieve with a 700 μm opening and were then air dried under a laminar flow hood for 2 min The arrangement of sclerotia on PDA and viability tests were the same as above described experiments. The data from the controls (0% NaOCl) were removed prior to analyzing the data. Resulting data were transformed using a binomial, where any growth of mycelia was recorded as 1, while no growth was recorded as 0. The experiment was repeated once.
Analysis of Data
Data variables containing binomial values were analyzed using Proc MIXED (SAS Institute, Ver. 9.1, Cary, NC). Means of binomial data are normally distributed (Steel and Torrie, 1960). Data from repeated experiments were combined when variances of trials were homogenous. For effect of sodium hypochlorite on bare sclerotia, fixed effects were the factorial of percent NaOCl, the amount of time in solution, deioinzed water rinse, and their interaction. Random effects were the two trials, replications within a trial, and the arrangement of PDA plates. For effect of sodium hypochlorite on sclerotia in soil, fixed effects were the factorial of soil type, percent NaOCl, and exposure time. Random effects were the two trials, replications within a trial, the interaction of trials by treatment, and replication by treatment within each trial was the residual error. Non-significant terms were removed from the full model until all terms remaining were significant at P ≤ 0.10. All removed terms were contained in a three-factor term in the random statement in the reduced model. Terms included in a significant three-factor term remained in the model regardless of significance rate of the single or two-factor terms. Pairwise comparisons of significant treatments or interactions in the reduced model were obtained with the PDIFF option, with a significance level of P = 0.05.
Results
Effect of Sodium Hypochlorite on Bare Sclerotia
Viability of sclerotia in the non-treated controls was 100% in each trial for both experiments. The interaction of NaOCl concentration and exposure time was significant (P < 0.0001). Sclerotia exposed to 0.6% NaOCl up to 10 min in solution germinated and grew in culture (Table 1). All sclerotia were killed at 10 min in a 3% NaOCl solution or at 5 min in a 6% NaOCl solution (Table 1). Sclerotia killed by the NaOCl treatment were spongy in texture and contained no visible traces of pigment. Rinsing sclerotia with sterile deionized water did not affect their survival.
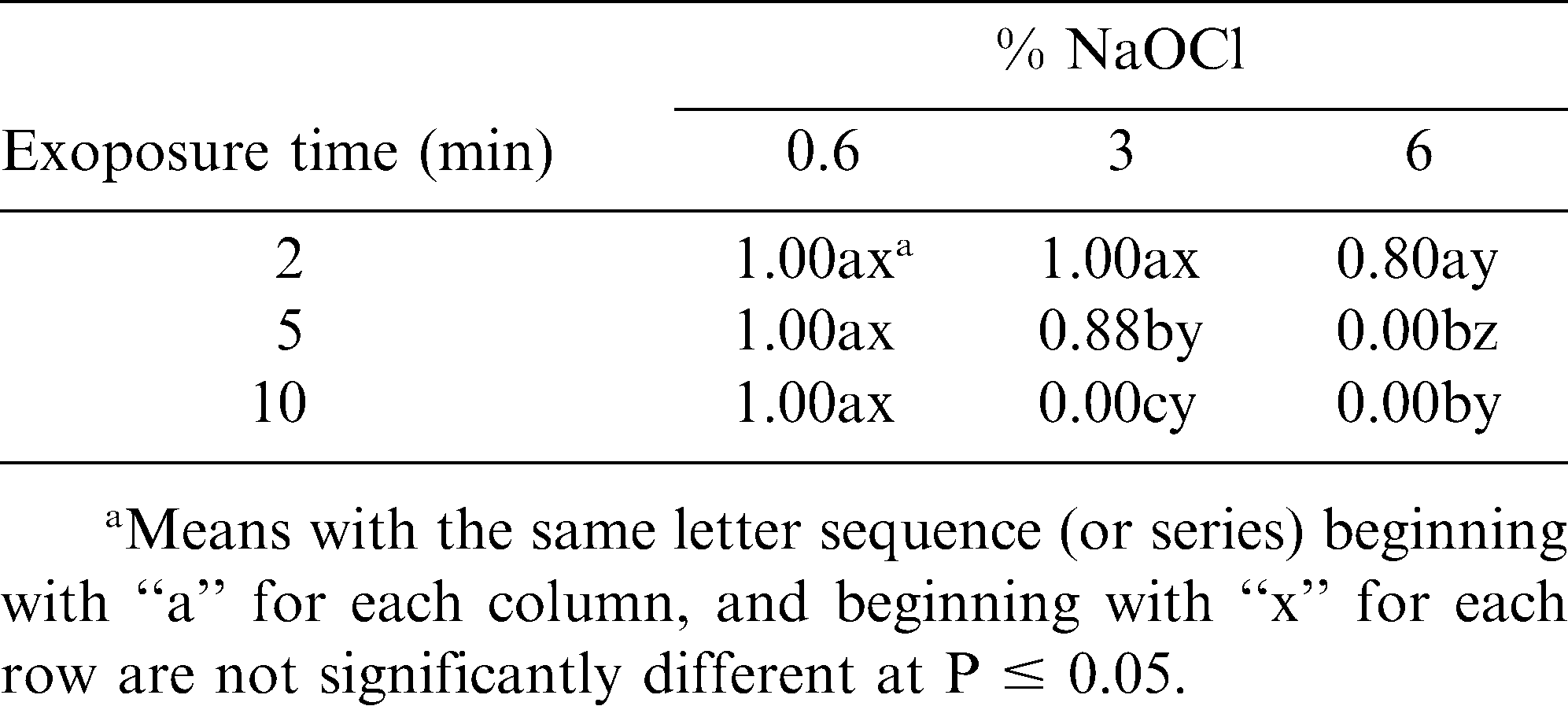
Effect of NaOCl concentration and exposure time on survival (1 = survival, 0 = death) of sclerotia of Sclerotinia minor produced in vitro.
Effect of Sodium Hypochlorite on Sclerotia in Soil
The interaction of NaOCl concentration, time in solution, and soil type was significant (P < 0.0001). At 3% NaOCl, soaking sclerotia up to 80 min in the sandy clay loam did not kill sclerotia (Table 2). Soaking sclerotia for either 60 or 80 min at the 6% concentration of NaOCl killed all sclerotia in both soil types (Table 2).
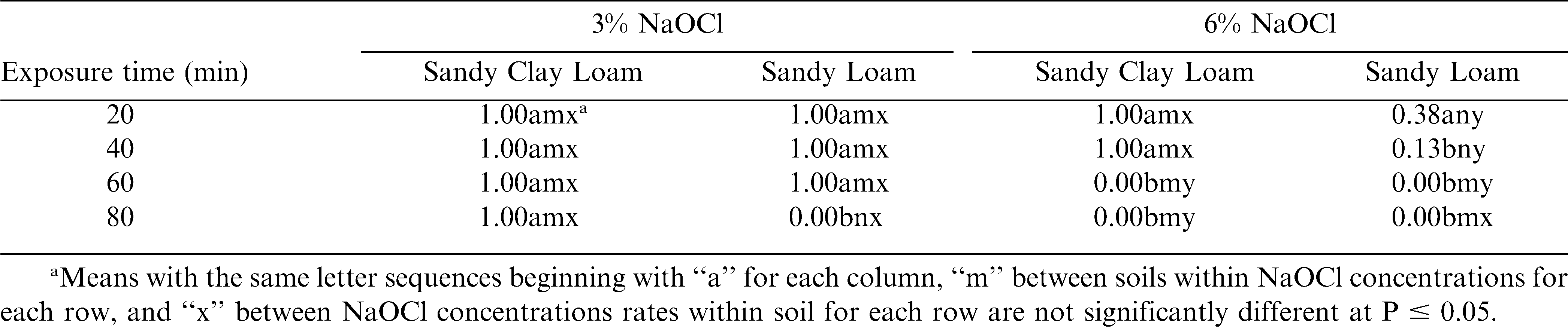
Effect of percent NaOCl, NaOCl exposure time, and soil type on survival (1 = survival, 0 = death) of sclerotia of Sclerotinia minor produced in vitro and mixed with two different field soils.
Discussion
Soaking in NaOCl at 6% for 1 hr destroyed the sclerotial rind of S. minor. However, soaking farm equipment with NaOCl at this concentration for 1 hr is impractical and would likely be corrosive to metal parts. Producers and field scouts concerned with transferring sclerotia from field to field should carefully clean equipment to remove all soil and plant debris. Effluent from contaminated equipment outside infested fields should be contained and allowed to dry. It may then be treated with 6% NaOCl for 1 hr to ensure sclerotia are killed, or simply returned to the already infested field.
Wymore and Lorbeer (1987) noted no change in germination when 0.5% NaOCl was applied to sclerotia of S. minor for up to 20 min. Studies with S. rolfsii have shown NaOCl can even increase mycelial germination of sclerotia (Linderman and Gilbert, 1972; Punja and Grogan, 1981; Punja et al., 1984). It was hypothesized that NaOCl affects rind cells by removing physical barriers that inhibit germination, and altering plasma membrane permeability (Punja and Grogan, 1981; Punja et al., 1984).
Previous research which determined the effects of various environmental parameters and chemical compounds on the germination of S. rolfsii sclerotia indicated that important physiological differences exist between rind cells of sclerotia produced on PDA verses field soil (Linderman and Glibert, 1972; Beute and Rodriguez-Kabana, 1979; Punja and Grogan, 1981; Punja et al., 1984). Punja and Grogan (1981) noted that sclerotia produced in culture had a thinner cortex and more medullary tissue. Physical differences in sclerotia of Sclerotinia spp. produced in the field versus on media have also been found (Le Tourneau, 1979; Merriman, 1976; Willetts and Wong, 1980; Wymore and Lorbeer, 1987). Sclerotia of Sclerotinia spp. produced on agar had less crude-fat, alcohol soluble material, and protein than those produced on peanut and bean (Phaseolus spp.) (Le Tourneau, 1979). Merriman (1976) found rinds of S. sclerotiorum (Lib.) de Bary sclerotia recovered from natural infestations were damaged due to microorganisms and did not survive in soil as long as those produced in culture. S. minor sclerotia produced under field conditions and unaffected by parasitic microorganisms may have a thicker rind and therefore may be even more difficult to destroy. Also, the effect of NaOCl on survivability of sclerotia embedded in crop residue and raw organic matter was not addressed. Therefore the exact times reported here for mortality of sclerotia produced in the lab may differ somewhat from that of sclerotia produced in the field, but the results should be generally applicable and relevant.
Acknowledgements
We appreciate the support of the Texas Peanut Producers and Texas AgriLife Research. Assistance in editing from Dr. Mark C. Black is greatly appreciated.
Literature Cited
Akem C. N. and Melouk H. A. 1990 Transmission of Sclerotinia minor in peanut from infected seed. Plant Dis 74: 216- 219.
Beute M. K. and Rodriguez-Kabana R. 1979 Effect of wetting and presence of peanut tissues on germination of sclerotia of Sclerotium rolfsii produced in soil. Phytopathology 69: 869- 872.
Le Tourneau D. 1979 Morphology, cytology, and physiology of Sclerotinia species in culture. Phytopathology 69: 887- 890.
Linderman R. G. and Gilbert R. G. 1972 Behavior of sclerotia of Sclerotium rolfsii produced in soil or in culture regarding germination stimulation of volatiles, fungistasis, and sodium hypochlorite treatment. Phytopathology 63: 500- 504.
Merriman P. R. 1976 Survival of sclerotia of Sclerotinia sclerotiorum in soil. Soil Biol. Biochem 8: 385- 389.
Porter D. M. and Beute M. K. 1974 Sclerotinia blight of peanuts. Phytopathology 64: 263- 264.
Punja Z. K. and Grogan R. G. 1981 Eruptive germination of sclerotia of Sclerotium rolfsii. Phytopathology 71: 1092- 1099.
Punja Z. K., Jenkin S. F., and Grogan R. G. 1984 Effect of volatile compounds, nutrients, and source of sclerotia on eruptive sclerotial germination of Sclerotium rolfsii. Phytopthology 74: 1290- 1295.
Wadsworth D. F. 1979 Sclerotinia blight of peanuts in Oklahoma and occurrence of the sexual stage of the pathogen. Peanut Sci 6: 77- 79.
Willetts H. J. and Wong J. A-L. 1980 The biology of Sclerotinia sclerotiorum, S. trifoliorum, and S. minor with emphasis on specific nomenclature. Bot. Rev 46: 101- 165.
Woodard K. E. and Simpson C. E. 1993 Characterization of growth and sclerotial production of Sclerotinia minor isolated from peanut in Texas. Plant Dis 77: 576- 579.
Wu B. M., Subbarao K. V., and Liu Y-B. 2008 Comparative survival of sclerotia of Sclerotinia minor and S. sclerotiorum. Phytopathology 98: 659- 665.
Wymore L. A. and Lorbeer J. W. 1987 Effect of cold treatment and drying on mycelial germination by sclerotia of Sclerotinia minor. Phytopathology 77: 851- 856.
Notes
Author Affiliations
1 Former Graduate Student, Professor, and Research Statistician, respectively. Texas AgriLife Research, 1102 East FM 1294, Lubbock, TX 79403. *Corresponding author: Present address - Dept. Soil and Crop Sciences, Texas A&M University, College Station, TX 77843. E-mail: jeffnw@hotmail.com