

Introduction
Peanut (Arachis hypogaea L.) is an important cash crop in the southeastern U.S. Peanut growers are demanding cultivars and management practices that reduce production costs so they can grow peanuts profitably to compete with imports. Peanut breeders have provided high yielding cultivars with good quality profiles for industry and consumer use. However, all of these cultivars are susceptible to one or more commonly occurring diseases. Few cultivars with multiple disease resistances are available, and none that has a sufficient level of leaf spot resistance to be grown without fungicide applications. Further, adverse weather conditions may delay timely sprays leading to heavy disease infestation that reduces yield and quality of peanuts. Thus, there is a need for high yielding, novel disease resistant peanut cultivars and progress in breeding for resistance has been slow due to the limited number of resistant genotypes available within the A. hypogaea germplasm. However, it is noteworthy that extensive work has been reported in developing leaf spot resistant runner cultivars using resistant plant introductions available within A. hypogaea gene pool (Branch, 2002; Gorbet and Shokes, 2002a and 2002b).
A potential source of disease resistance genes for peanut improvement is the wild species. The genus contains about 80 diploid (2n = 2x = 20), aneuploid (2n = 2x = 18) and tetraploid (2n = 4x = 40) species that have been grouped into nine botanical sections (Krapovickas and Gregory, 1994; Valls and Simpson, 2005) and A. hypogaea will only hybridize with species in section Arachis. Disease evaluations of section Arachis species revealed several with high levels of resistance to many important pathogens and pests (Lynch and Mack, 1995; Stalker and Moss, 1987; Stalker and Simpson, 1995). Although wild species of section Arachis have been suggested as sources of resistance genes since the 1960s (Smartt and Gregory, 1967), only two disease resistant, runner commercial cultivars have been released to-date from interspecific hybridization (Simpson and Starr, 2001; Simpson et al., 2003). A major reason for this slow and limited use of Arachis species for peanut improvement has been their ploidy level differences and genomic incompatibilities with A. hypogaea (Tallury et al., 2005). Nonetheless, several tetraploid interspecific hybrid lines derived from A. cardenasii Krapov. & W.C. Gregory, A. stenosperma Krapov. & W.C. Gregory, A. diogoi Hoehne, A. batizocoi Krapov. & W.C. Gregory and A. correntina (Burkart) Krapov. & W.C. Gregory, have been developed in the NCSU peanut genetics program and 16 germplasm lines were released (Stalker and Beute, 1993; Stalker and Lynch, 2002; Stalker et al., 2002a; Stalker et al., 2002b; Isleib et al., 2006a). These germplasm lines are publicly available and have high levels of resistances to early and late leaf spots (Cercospora arachidicola Hori and Cercosporidium personatum (Berk. & Curt.) Deighton), root-knot nematode (Meloidogyne arenaria [Neal] Chitwood), tomato spotted wilt virus (TSWV), Sclerotinia blight (S. minor Jagger), Cylindrocladium black rot (CBR, C. parasiticum Crous, Wingfield & Alfenas), web blotch (Phoma arachidicola) and several insects, such as potato leafhopper (Empoasca fabae [Harris]), corn earworm (Helicoverpa zea [Boddie]), and lesser cornstalk borer (Elasmopalpus lignosellus [Zeller]). Several of these germplasm lines and other disease resistant selections have been hybridized with susceptible commercial cultivars and additional selections from the resulting progenies were evaluated in the present study (Table 1) for possible cultivar releases.
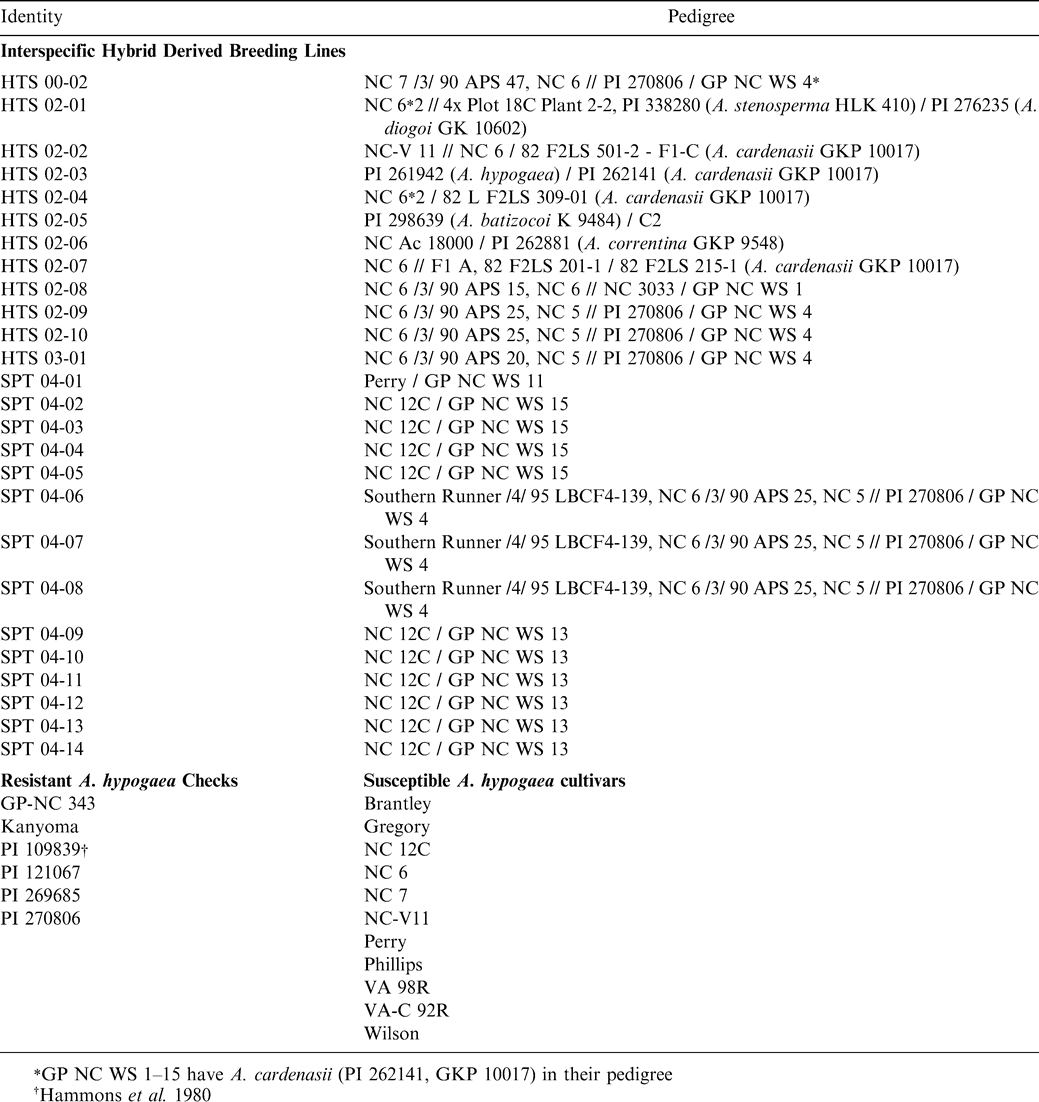
Experimental material with their genetic background.
The overall goal of our research program is to develop peanut cultivars with resistance to the most common diseases in the V-C production area, which include early leaf spot (ELS), TSWV, Sclerotinia blight (SB) and CBR. The specific goals of the research reported in this paper are 1) to compare tetraploid, interspecific hybrid derived breeding lines with commercial peanut cultivars for their agronomic performance and leaf spot resistance, and 2) to evaluate the potential of tetraploid interspecific hybrid derived breeding lines for commercial cultivation in the V-C peanut production area.
Materials and Methods
Twenty-six tetraploid, interspecific hybrid derived breeding lines, six A. hypogaea lines resistant to early- and late leaf spots (resistant checks), and 11 commercial cultivars (susceptible checks) (Table 1) [NC 6 (Campbell et al., 1977); NC 12C (Isleib et al., 1997); Gregory (Isleib et al., 1999); Perry (Isleib et al., 2003); Brantley ( Isleib et al., 2006b); Phillips ( Isleib et al., 2006c); VA-C 92R (Mozingo et al., 1994); VA 98R (Mozingo et al., 2000); Wilson (Mozingo et al., 2004); NC 7 (Wynne et al., 1979) and NC-V11 (Wynne et al., 1991)] were evaluated in field tests at the Peanut Belt Research Station in Lewiston, NC from 2004 to 2006. Experimental plots consisted of 2-rows each, 7.3 m long on 91 cm wide beds and 28 seeds/row were planted with 25 cm spacing. The plots were planted on 11 May in each of the 3 years. In each year the experimental design was a lattice with two replications, with a 10×10 simple square lattice in 2004 and 10×9 double rectangular lattices in 2005. The additional lines evaluated in these experiments (57 in 2004, 47 in 2005 and 2006) were experimental A. hypogaea lines from the N.C. State Univ. peanut-breeding project. Data on these experimental lines were not reported in this study (T.G. Isleib, unpublished data). Recommended management practices were followed during the growing of the crop but no fungicidal sprays were used to control leaf spots.
Defoliation ratings were recorded on plots on 2, 14, and 21 Sept. in 2004, on 21 and 27 Sept. in 2005, and on 9 and 21 Sept. in 2006, respectively. The severity of the disease was rated on a proportional scale from 1 = no defoliation (resistant) to 9 = complete defoliation (susceptible). The 3 yrs of data were combined to generate mean disease scores. Pod yield and grade were measured on all plots and combined to generate means. Analysis of variance was performed on defoliation scores, yields and grades. Means were adjusted for block effects in the incomplete block design if block effects were found to be significant by F-test. The means for the 43 entries from each year were used to perform an analysis of variance as if for a randomized complete block design using years as the blocking variable. Means were separated by t-tests. All analyses were performed using the general linear models procedure (PROC GLM) of the SAS statistical software package (SAS Institute, Inc. 2001).
Results and Discussion
Leaf spot evaluations
Significant variation among the 43 entries was observed (P<0.05) for both defoliation score and yield (Table 2). In 2004, the experimental plots showed higher incidence of ELS than in the other 2 yrs. Higher incidence of LLS was observed in the experimental plots in 2005 at Lewiston and both leaf spots were present in 2006. Although ELS has been the major disease in the V-C production area, LLS also causes significant damage as was observed in 2005. Together, these two diseases can result in up to 50% yield losses to growers (Shew et al., 1995). Cultivars with resistance to both diseases will greatly benefit the V-C peanut growers. As a group, the mean defoliation score of the cultivars was 6.8-±0.1 whereas the corresponding value for the resistant checks was 4.3±0.2 and for the interspecific hybrid breeding lines was 5.3±0.1, respectively. Thus, significant differences were observed for mean defoliation score (Table 2). Among the cultivars grown in the V-C area, the mean defoliation score ranged from 6.4 for the obsolete cultivar NC 6 to 7.4 for Wilson (Table 2). In the resistant A. hypogaea checks, the mean defoliation score varied from 3.0 in PI 269685 to 5.5 in Kanyoma. Correspondingly, the mean defoliation scores of the 26 interspecific hybrid derived breeding lines ranged from 4.4 in SPT 04-01 to 6.3 in SPT 04-13 and SPT 04-14. These three lines were derived from A. cardenasii. The line, HTS 02-01, derived from an amphidiploid of A. stenosperma × A. diogoi, had a mean defoliation score of 5.1, whereas, the A. batizocoi (HTS 02-05) and A. correntina (HTS 02-06) derivatives had mean defoliation scores of 5.9 and 5.0, respectively. Nine of the 26 interspecific hybrid breeding lines had mean defoliation scores of 5 or lower whereas the lowest mean defoliation score among the cultivars was a 6.4 for NC 6. Although a difference of 1.4 points or more in defoliation scores between the cultivars and the interspecific hybrid derived breeding lines may not seem significant, nevertheless, the leaf spot resistance levels observed in the interspecific hybrid derived breeding lines warrant further field evaluations for releases. In a study involving the components of resistance to late leaf spot and rust among 15 interspecific hybrid derivatives, Dwivedi et al. (2002) indicated that the disease score, which is primarily based on percentage defoliation, integrates all components of resistance and an optimum combination of these components results in lower defoliation scores. Thus, because of their lower defoliation scores, the interspecific hybrid derived breeding lines in this study may have accumulated an optimum combination of different components of resistance.
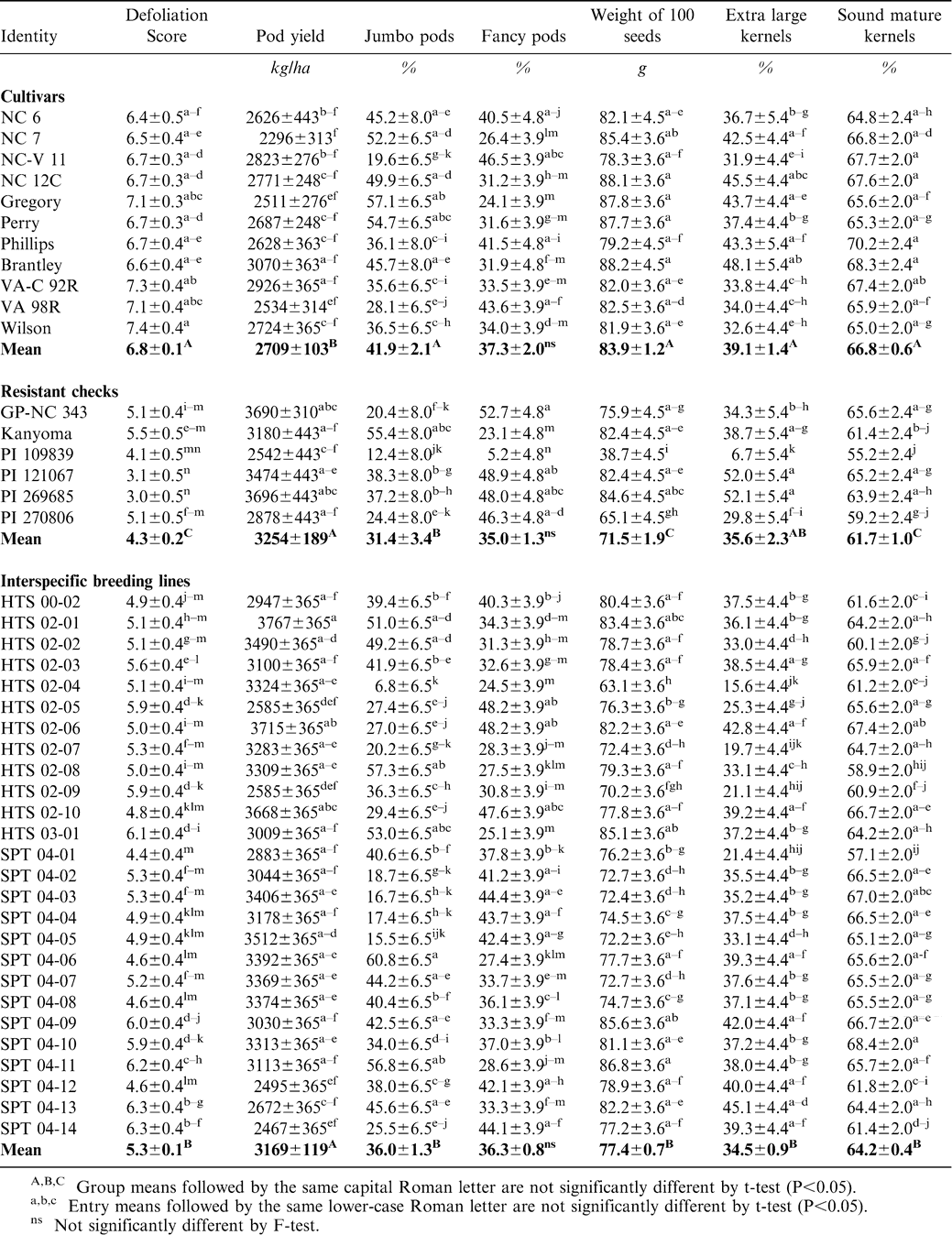
Means of defoliation score, yield, and selected grade characteristics of susceptible cultivars, resistant A. hypogaea checks and tetraploid interspecific hybrid derived breeding lines evaluated at the Peanut Belt Res. Stn., Lewiston, NC from 2004–2006.
Currently, peanut growers use foliar application of fungicides to control leaf spots, usually beginning in early July and continuing until mid-Sept following a 14-d spray schedule or a weather based spray advisory. This requires five to six sprays during the crop season to control leaf spots with chemical costs of$371–494/ha. The interspecific hybrid derived breeding lines have high levels of leaf spot resistance even under heavy disease pressure, unlike some of the moderately leaf spot resistant V-C peanut cultivars which express severe defoliation symptoms under heavy disease pressure, and are a promising source for the development of leaf spot resistant cultivars for the V-C area.
Yield Evaluations
Although disease-resistant cultivars may reduce fungicidal sprays and lower production costs, the most important traits to a grower are the yield and grade because they determine the price and eventual profits. Large-seeded virginia type cultivars, such as Gregory, Perry, NC 12C, NC-V11, VA 98R and Wilson, dominate the V-C production area. In field tests conducted without chemical control of leaf spots, the mean yields ranged from 2296±313 kg/ha in NC 7 to 3070±363 kg/ha in Brantley, whereas 19 of the 26 interspecific hybrid derived breeding lines produced at least 3000 kg/ha without chemical control of leaf spots (Table 2). Of these, the highest yielding line, HTS 02-01 which is derived from the amphidiploid, [A. hypogaea × 4x (A. stenosperma × A. diogoi)] yielded 3767±365 kg/ha. Thus, the interspecific derived breeding lines yielded, on average, at least 300 kg more than the existing commercial cultivars with no fungicidal sprays (Table 2). As a result, a grower should be able to reduce or possibly eliminate the number of fungicidal sprays to control leaf spots and substantially lower production costs by reducing fuel and chemical costs.
Grade and Flavor Evaluations
Overall, the means of the selected grade characteristics of the interspecific breeding lines were slightly inferior to those of the cultivars (Table 2). For example, in the present study, Brantley had mean ELK content of 48.1% and SMK content of 68.3%. Among the interspecific hybrid derived breeding lines, the corresponding values for SPT 04-13 were 45.1 and 64.4%. Overall, 22 of the 26 interspecific hybrid derived breeding lines had ELK and SMK values similar to those of the cultivars (Table 2). Some of these lines exhibited values for jumbo pods, fancy pods and 100-seed weight similar to those of the cultivars (Table 2). Sixteen of the 26 interspecific hybrid derived breeding lines and NC 7, the commercial flavor standard for V-C area, were also evaluated for sensory quality. No significant variation among test entries and NC 7 was found for the roasted peanut, sweet, or bitter sensory attributes (Tallury et al., 2008) indicating that the interspecific hybrid derived breeding lines did not exhibit flavor degradation, an important attribute if these lines are to be considered for cultivar release in the V-C area.
Conclusions
Currently grown cultivars in the V-C area produced less yield with more disease than the interspecific hybrid derived breeding lines. Many interspecific hybrid derived breeding lines yielded 3000 kg/ha without chemical control of leaf spots. This indicates that the interspecific hybrid derived breeding lines evaluated in this study have superior leaf spot resistance combined with high yield. Because the diploid Arachis species used in the pedigrees of the interspecific hybrid breeding lines have a broad range of disease resistances to several common pathogens and pests of peanut (Stalker and Moss, 1987), they may also be resistant to other diseases of peanut. These lines need to be evaluated in multiple environments in the V-C area before a recommendation for commercial use can be made.
Acknowledgements
This research was supported by funds from NC Peanut Growers Association, NC Crop Improvement Association and NC Foundation Seed Producers, Inc. Susan Copeland and Philip Rice helped with field planting and recording data.
Literature Cited
Branch W. D. 2002 Registration of ‘Georgia-01R’ peanut. Crop Sci 42 : 1750 – 1751 .
Campbell W. V. , Emery D. A. , and Gregory W. C. 1971 Registration of GP-NC 343 peanut germplasm (Reg. No. GP 1). Crop Sci 11 – 605 .
Campbell W. V. , Wynne J. C. , Emery D. A. , and Mozingo R. W. 1977 Registration of NC 6 peanuts (Reg. No. 20). Crop Sci 17 : 346 .
Dwivedi S. L. , Pande S. , Narayan Rao J. , and Nigam S. N. 2002 Components of resistance to late leaf spot and rust among interspecific derivatives and their significance in a foliar disease resistance breeding in groundnut (Arachis hypogaea L.). Euphytica 125 : 81 – 88 .
Gorbet D. W. and Shokes F. M. 2002a Registration of ‘C-99R’ peanut. Crop Sci 42 : 2207 .
Gorbet D. W. and Shokes F. M. 2002b Registration of ‘Florida MDR 98’ peanut. Crop Sci 42 : 2207 – 2208 .
Hammons R. O. , Sowell G. , and Smith D. H. 1980 Registration of Cercospora arachidicola-resistant peanut germplasm (Reg. No. GP 10; PI 109839). Crop Sci 20 : 292 .
Isleib T. G. , Rice P. W. , Bailey J. E. , Mozingo R. W. , and Pattee H. E. 1997 Registration of ‘NC 12C’ peanut. Crop Sci 37 : 1976 .
Isleib T. G. , Rice P. W. , Mozingo R. W. , Bailey J. E. , Mozingo R. W. , and Pattee H. E. 2003 Registration of ‘Perry’ peanut. Crop Sci 43 : 739 – 740 .
Isleib T. G. , Rice P. W. , Mozingo R. W. , Copeland S. C. , Graeber J. B. , Shew B. B. , Smith D. L. , Melouk H. A. , and Stalker H. T. 2006a Registration of N96076L peanut germplasm. Crop Sci 46 : 2329 – 2330 .
Isleib T. G. , Rice P. W. , Mozingo R. W. , Copeland S. C. , Graeber J. B. , Novitzky W. P. , Pattee H. E. , Sanders T. H. , Mozingo R. W. , and Coker D. L. 2006b Registration of ‘Brantley’ peanut. Crop Sci 46 : 2309 – 2311 .
Isleib T. G. , Rice P. W. , Mozingo R. W. , Copeland S. C. , Graeber J. B. , Pattee H. E. , Sanders T. H. , Mozingo R. W. , and Coker D. L. 2006c Registration of ‘Phillips’ peanut. Crop Sci 46 : 2308 – 2309 .
Isleib T. G. , Rice P. W. , Mozingo R. W. , Mozingo R. W. , and Pattee H. E. 1999 Registration of ‘Gregory’ peanut. Crop Sci 39 : 1526 .
Krapovickas A. and Gregory W. C. 1994 Taxonomy of the genus Arachis (Leguminosae). Bonplandia 8 : 1 – 186 .
Lynch R. E. and Mack T. P. 1995 Biological and biotechnical advances for insect management in peanut. 95 – 159 In Pattee H. E. and Stalker H. T. Advances in Peanut Science Amer. Peanut Res. Educ. Soc Stillwater, OK .
Mozingo R. W. , Coffelt T. A. , and Isleib T. G. 2000 Registration of VA 98R peanut. Crop Sci 40 : 1202 – 1203 .
Mozingo R. W. , Coffelt T. A. , Swann C. W. , and Phipps P. M. 2004 Registration of ‘Wilson’ peanut. Crop Sci 44 : 1017 – 1018 .
Mozingo R. W. , Wynne J. C. , Porter D. M. , Coffelt T. A. , and Isleib T. G. 1994 Registration of VA-C 92R peanut. Crop Sci 34 : 539 – 540 .
SAS Institute, Inc 2001 SAS/STAT Software: Changes and Enhancements, Release 8.2 SAS Institute, Inc Cary, NC 332 .
Shew B. B. , Beute M. K. , and Stalker H. T. 1995 Toward sustainable peanut production: Progress in breeding for resistance to foliar and soil borne pathogens of peanut. Plant Disease 79 : 1259 – 1261 .
Simpson C. E. and Starr J. L. 2001 Registration of ‘COAN’ peanut. Crop Sci 41 : 918 .
Simpson C. E. , Starr J. L. , Church G. T. , Burow M. D. , and Paterson A. H. 2003 Registration of ‘NemaTAM’ peanut. Crop Sci 43 : 1561 .
Smartt J. and Gregory W. C. 1967 Interspecific cross-compatibility between the cultivated peanut Arachis hypogaea L. and other members of the genus Arachis. Oléagineux 22 : 455 – 459 .
Stalker H. T. and Beute M. K. 1993 Registration of four interspecific peanut germplasm lines resistant to Cercospora arachidicola. Crop Sci 33 : 1117 .
Stalker H. T. , Beute M. K. , Shew B. B. , and Barker K. R. 2002a Registration of two root-knot nematode-resistant peanut germplasm lines. Crop Sci 42 : 312 – 313 .
Stalker H. T. , Beute M. K. , Shew B. B. , and Isleib T. G. 2002b Registration of five leafspot-resistant peanut germplasm lines. Crop Sci 42 : 314 – 316 .
Stalker H. T. and Lynch R. E. 2002 Registration of four insect-resistant peanut germplasm lines. Crop Sci 42 : 313 – 314 .
Stalker H. T. and Moss J. P. 1987 Speciation, cytogenetics, and utilization of Arachis species. Adv. Agronomy 41 : 1 – 40 .
Stalker H. T.
and
Simpson C. E.
1995
Genetic resources in Arachis.
14
–
53
In
Pattee H. E.
and
Stalker H. T.
Advances in Peanut Science
Amer. Peanut Res. Educ. Soc
Stillwater, OK
.
Tallury S. P. , Hilu K. W. , Milla S. R. , Friend S. A. , Alsaghir M. , Stalker H. T. , and Quandt D. 2005 Genomic affinities in Arachis section Arachis (Fabaceae): Molecular and cytogenetic evidence. Theor. Appl. Genet 111 : 1229 – 1237 .
Tallury S. P. , Pattee H. E. , Isleib T. G. , and Stalker H. T. 2008 Flavor profiles of wild species-derived peanut breeding lines. Peanut Sci 35 : 81 – 85 .
Valls J. F. M. and Simpson C. E. 2005 New species of Arachis from Brazil, Paraguay, and Bolivia. Bonplandia 14 : 1 – 2 .
Wynne J. C. , Coffelt T. A. , Mozingo R. W. , and Anderson W. F. 1991 Registration of ‘NC-V11’ peanut. Crop Sci 31 : 484 – 485 .
Wynne J. C. , Mozingo R. W. , and Emery D. A. 1979 Registration of NC 7 peanut (Reg. No. 22). Crop Sci 19 : 563 .
Notes
1Dept. Crop Science, Box 7629, N. C. State Univ., Raleigh, NC 27695-7629.
*Corresponding author (E-mail:tallury@ncsu.edu)