

Introduction
Throughout the world, aflatoxin contamination is considered one of the most serious food and feed safety issues. Chronic problems with aflatoxin contamination occur in the southern U.S., and are particularly troublesome in peanut, corn, cottonseed, and tree nuts. The impact of aflatoxin contamination on the agricultural economy is especially devastating during drought years when aflatoxin affects the more northern areas including the Midwestern corn belt. Economic losses in the years of major aflatoxin outbreaks have been estimated in the hundreds of millions of dollars. The realization of the unique nature of the aflatoxin problem and the need for novel technologies to ameliorate the impact of this problem became a focal point of discussion in 1988 at the first U.S. Aflatoxin Elimination Workshop held in New Orleans, LA. The Annual Aflatoxin Workshops (Table 1) have served as a forum to assemble USDA-ARS scientists, university faculty, representatives of the different commodities and industries, and international participants in a unique cooperative effort to develop aflatoxin control strategies through research and development.
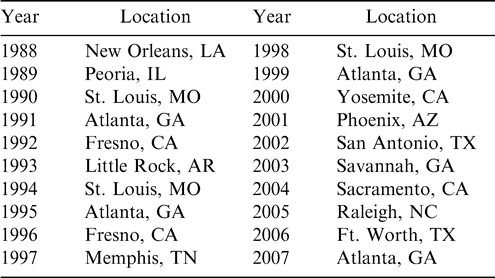
USDA Annual Multicrop Aflatoxin Elimination Workshop.
Peanut (Arachis hypogaea L.), also called groundnut, is an important staple crop in many areas of the world and also is one of the most susceptible host species for Aspergillus flavus Link:Fr., which is the causal organism producing aflatoxin. The International Conference on Groundnut Aflatoxin Management and Genomics was held 5-9 November, 2006, in Guangzhou, China, and provided an international forum for scientific presentations and discussions about the latest accomplishments, strategies and cooperation in the prevention of aflatoxin contamination through genetics and genomics. Our presentation at this conference focused on searching for strategies in prevention of preharvest aflatoxin contamination through understanding the mechanisms of aflatoxin formation, pathogenicity of the fungus, and host crop resistance along with recent research on crop-fungus interaction using microarray by gene expression profilings. The joint effort between USDA-ARS at Tifton, GA and New Orleans, LA, and the J. Craig Venter Institute (JCVI) to produce a microarray to study the interaction between Aspergillus and peanut is also reviewed in this paper.
Aflatoxins and Importance of Aflatoxin Contamination
Aflatoxins
Mycotoxins are low molecular weight secondary metabolites produced by filamentous fungi and induce various degrees of toxicity to vertebrates, invertebrates, plants, and microorganisms. Aflatoxin was first associated with an outbreak of “Turkey X disease”, which occurred in 1960 near London, England, and killed approximately 100,000 poults (Blount, 1961; Forgacs and Carll, 1962). The cause of the disease was later associated with the feeding of peanut (groundnut) meal infested by A. flavus, and thus the toxins were named “aflatoxin” for the A. flavus toxin.
Attention has been given to the occurrence, biosynthesis, and toxicity of aflatoxins since these compounds are both carcinogenic and the most toxic of the known mycotoxins (Cole and Cole, 1987; Van Egmond, 1989; Yu et al., 2004a, b,c; Yu et al., 2006). Aflatoxins belong to a family of compounds with difuranocoumarins. Aflatoxins B1, B2, G1, and G2 (AFB1, AFB2, AFG1, and AFG2) are the four major aflatoxins based on their blue (B) or green (G) fluorescence under ultraviolet light and their relative mobility by thin-layer chromatography on silica gel. Aflatoxin M1 is a hydroxylated derivative metabolized from aflatoxin B1 by cows and secreted in milk (Van Egmond, 1989). In addition to aflatoxins B1 and B2, A. flavus produces cyclopiazonic acid, kojic acid, beta-nitropropionic acid, aspertoxin, aflatrem and aspergillic acid (Goto et al., 1996). Aspergillus parasiticus Speare produces G1 and G2 in addition to aflatoxins B1 and B2.
Food safety and economic impact
Aspergillus flavus is a weak opportunistic plant pathogen and secretes aflatoxins into many agricultural crops. Preharvest aflatoxin contamination can occur when A. flavus infects peanut pods, corn ears, and cotton bolls with insect or mechanical damage (or if tissues are not amaged). Postharvest aflatoxin contamination can be problematic if grain storage is poorly managed. Outbreaks of acute aflatoxicosis from contaminated food in humans have been documented in Kenya, India, Malaysia, and Thailand as reported by the Council for Agriculture Science and Technology (CAST, 2003). For example, an outbreak of severe aflatoxicosis in humans occurred in more than 150 villages in western India in 1974 where 397 persons were affected and 108 persons died (Krishnamachari et al., 1975). The largest and most severe outbreak of acute aflatoxicosis documented worldwide occurred in Kenya during 2004 and involved 317 cases and 125 deaths, mainly among children, due to consumption of aflatoxin contaminated corn (Lewis et al., 2005; CDC, 2004).
Low dose consumption of aflatoxin contaminated food stuff causes chronic aflatoxicosis resulting in cancer, suppression of immunological reponses, and other “slow” pathological conditions. The liver is the primary target organ by toxic and carcinogenic aflatoxins. Cytochrome P450 enzymes in the liver convert aflatoxins to the reactive 8,9-epoxide form, which is capable of binding to both DNA and proteins (Eaton and Groopman, 1994). Aflatoxin B1-DNA adducts can result in the GC to TA transversions in the p53, a DNA-repair, tumor-suppressor gene, at codon 249. Inactivation of the p53 tumor suppressor gene leads to the development of primary liver cancer (Bressac et al., 1991; Hsu et al., 1991).
Aflatoxin contamination also has a significant economic impact on worldwide agriculture. In the developing countries, food safety is the major problem where detection and decontamination policies are impractical. Due to food shortage in those countries, routine consumption of aflatoxin-contaminated food is widespread. The liver cancer incidence rates are 2 to 10× higher in developing countries than in developed countries (Henry et al., 1999). In developed countries, the maximum allowable amount of aflatoxin in food and feed for human consumption and for livestock has been mandated by law. A guideline of 20 ppb (parts per billion) aflatoxin in food or feed substrate is the maximum allowable limit imposed by the U.S. Food and Drug Administration. The European Union has a maximum level of 2 ppb for aflatoxin B1 and 4 ppb for total aflatoxins. The aflatoxin contaminated commodities are often destroyed if the aflatoxin content is higher than the mandated levels. This results in billions of dollars in yearly losses worldwide. In some parts of the southern U.S. such as in the southeast peanut, southern cotton belt, and mid-south corn farming regions, severe outbreaks of aflatoxin contamination occur frequently and resulted in enormous economic losses (CAST, 1989; Robens and Cardwell, 2005).
Genetics and Biochemistry of Aflatoxin Biosynthesis
Aflatoxin biosynthetic pathway, genes and enzymes
The elucidation of the aflatoxin biosynthetic pathway has been one area of research focus during the last decade. The major biochemical pathway steps have been determined and the chemical structures of aflatoxin intermediates have been characterized (Minto and Townsend, 1997; Payne and Brown, 1998; Yu, 2004; Yu et al., 2004a, c; Yu et al., 2005). At least 23 enzymatic reactions are involved in aflatoxin formation. No less than 15 structurally-defined aflatoxin intermediates have been identified in the aflatoxin/ST (sterigmatocystin) biosynthetic pathway.
Aflatoxins are synthesized from malonyl CoA, first with the formation of hexanoyl CoA, followed by formation of a decaketide anthraquinone (Bhatnagar et al., 1992; Minto and Townsend, 1997). There are two fatty acid synthases (FAS) and a polyketide synthase (PKS) involved in the synthesis of the polyketide from acetyl CoA (Watanabe and Townsend, 2002). Norsolorinic acid (NOR) is the first stable aflatoxin intermediate identified in the pathway (Bennett, 1981). Aflatoxins are formed after a series of oxidation-reduction reactions. The generally accepted aflatoxin biosynthetic pathway scheme is: a hexanoyl CoA precursor → norsolorinic acid, NOR → averantin, AVN → hydroxyaverantin, HAVN → Oxoaverantin, OAVN → averufin, AVF → hydroxyversicolorone, HVN → versiconal hemiacetal acetate, VHA → versiconal, VAL → versicolorin B, VERB → versicolorin A, VERA → demethyl-sterigmatocystin, DMST → sterigmatocystin, ST → O-methylsterigmatocystin, OMST → aflatoxin B1 and aflatoxin G1 (Fig. 1). After the VHA step, there is a branch point in the pathway that leads to aflatoxins B2 and G2 formation (Yabe et al., 2003; Yu et al., 1998; Yu et al., 2004c).
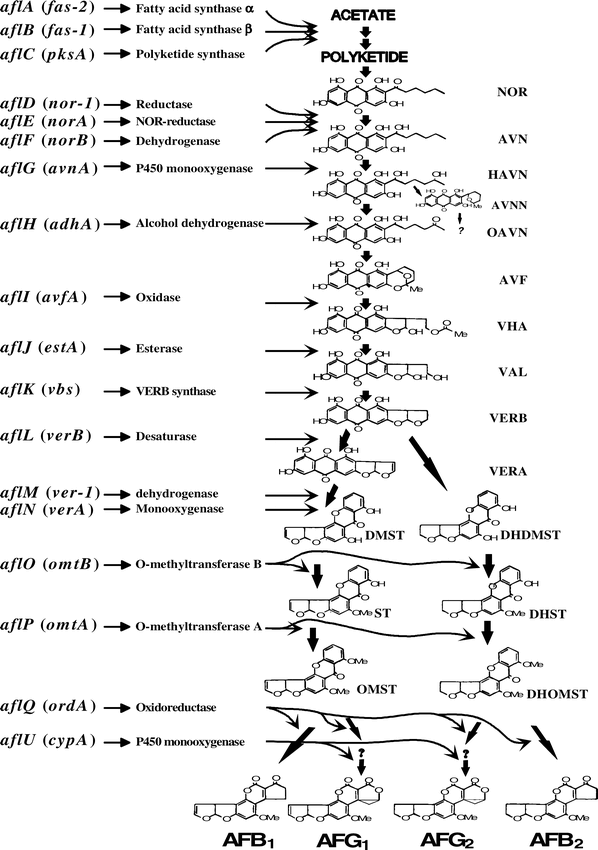
A total of 29 genes or open reading frame (ORF) are involved in aflatoxin formation. The first aflatoxin pathway gene identified was aflD (nor-1) that encodes a ketoreductase in A. parasiticus (Chang et al., 1992) for the conversion of norsolorinic acid (NOR) to averantin (AVN). Disruption or deletion of the aflD (nor-1) gene leads to the accumulation of a brick-red pigment in the hyphae and blocks the synthesis of all aflatoxins and their intermediates beyond NOR (Bennett, 1981). A gene named aflR present in A. parasiticus and A. flavus, as well as in A. nidulans Eidam (originally named afl-2 and apa-2), has been cloned by Chang et al., (1993) and Payne et al. (1993). This is a positive regulatory gene involved in both the aflatoxin pathway gene expression in A. flavus and A. parasiticus and sterigmatocystin (ST) pathway gene expression in A. nidulans. These four genes for aflatoxin or ST biosynthesis are milestone discoveries.
There are two separate pathways leading to B-Group (AFB1 and AFB2) and G-Group (AFG1 and AFG2) aflatoxins (Yabe et al., 1988). A gene, named aflQ (ordA), encoding a cytochrome P-450 monooxygenase, was demonstrated to be responsible for the conversion of O-methylsterigmatocystin (OMST) to AFB1 and AFG1, and demethyldihydrosterigmatocystin (DMDHST) to AFB2 and AFG2 (Prieto and Woloshuk, 1997; Yu et al., 1998) in A. parasiticus and A. flavus. Expression and substrate feeding, using a yeast system, demonstrated that an additional enzyme was required for formation of G-group aflatoxins (AFG1 and AFG2) (Yu et al., 1998). Functional studies demonstrated that the aflU (cypA) gene in A. parasiticus, encoding a cytochrome P450 monooxygenase, is responsible for the conversion of OMST to AFG1 and DHOMST to AFG2 (Ehrlich et al., 2004). A partial deletion of this gene results in loss of G-group aflatoxin production in A. flavus (Ehrlich et al., 2004).
The observation that the aflD (nor-1) and aflM (ver-1) genes were linked with the regulatory gene aflR in a common cosmid clone was the initial evidence indicating that aflatoxin pathway genes were clustered (Skory et al., 1993; Trail et al., 1995b). The completed aflatoxin pathway gene cluster was established when an 82 kb DNA sequence harboring a total of 29 aflatoxin biosynthetic pathway genes (or ORFs) and four sugar utilization genes were reported (Yu et al., 2004c). The primary advantage of gene clustering may be for the purpose of coordinated gene expression.
Factors affecting aflatoxin biosynthesis
Biotic and abiotic factors, either nutritional or environmental, are known to affect aflatoxin production in toxigenic Aspergillus species, although the molecular mechanisms for these effects are still unclear (Payne and Brown, 1998; Guo et al., 2005a, b). Nutritional factors such as carbon, nitrogen, amino acid, lipid, and trace elements affect aflatoxin production (Feng and Leonard, 1998; Payne and Brown, 1998; Cuero et al., 2003). Simple sugars such as glucose, sucrose, and maltose support aflatoxin formation, but peptone, sorbose, or lactose do not promote the toxin (Payne and Brown, 1998). However, the mechanism by which a carbon source is involved in the regulation of aflatoxin pathway gene expression is poorly understood. Nitrogen source affects aflatoxin formation in varying ways and production levels are different when Aspergillus spp. is on a nitrate versus a nitrite medium (Payne and Brown, 1998). Certain amino acids also can have opposing effects on aflatoxin production (Payne and Hagler, 1983). Recent studies using A. flavus show that tryptophan inhibits aflatoxin formation while tyrosine spikes aflatoxin production (Wilkinson et al., 2007). Micronutrients (metal ions) were also reported to affect aflatoxin pathway gene expression (Bennett et al., 1979; Cuero et al., 2003). Lipids have tremendous effects on aflatoxin formation, not only as a nutritive source but also as substrates metabolized for acyl-CoA starter units (Maggio-Hall et al., 2005) and as signaling molecules (Brodhagen and Keller, 2006).
Temperature, pH, water activity (drought stress) and other stresses are external environmental factors that can affect aflatoxin production (Cotty, 1988; Payne and Brown, 1998; Guo et al., 2005b; Kim et al., 2005, 2006; Sobolev et al., 2007). Studies suggest that aflR transcription is responsive to a G-protein signaling cascade that is mediated by protein kinase A (Hicks et al., 1997). This signaling pathway may respond to the environmental effects on aflatoxin biosynthesis. Optimal aflatoxin production is observed at temperatures near 30C (28 to 35C). Aflatoxin production is closely related to pH changes where biosynthesis in A. flavus occurs in acidic media, but is inhibited in alkaline media (Cotty, 1988).
Fungal developmental processes are associated with secondary metabolism such as sporulation and sclerotial formation (Bennett et al., 1986; Calvo et al., 2002; Chang et al., 2002; Yu, 2006). Similar environmental conditions required for secondary metabolism and for sporulation are observed. It was also reported that the spore formation and secondary metabolite formation occur at about the same time (Hicks et al., 1997; Trail et al., 1995a). Mutants that have suppressed sporulation were unable to produce aflatoxins (Bennett and Papa, 1988). Some compounds in A. parasiticus that inhibit sporulation also have been shown to inhibit aflatoxin formation (Reib, 1982).
Oxidative stress induced aflatoxin biosynthesis in A. parasiticus has long been reported. Kim et al. (2006) showed that treatment of A. flavus with tert-butyl hydroperoxide induced significant increases in aflatoxin production. Similar treatment of A. parasiticus also induced aflatoxin production (Reverberi et al., 2005; Reverberi et al., 2006). However, hydrolysable tannins significantly inhibit aflatoxin biosynthesis with the main antiaflatoxigenic constituent being gallic acid (Mahoney and Molyneux, 2004). Gallic acid reduces expression of genes within the aflatoxin biosynthetic cluster. When certain phenolics or antioxidants such as ascorbic acid are added to oxidatively stressed A. flavus, aflatoxin production significantly declines with no effect on fungal growth (Kim et al. 2006).
Functional Genomics and Control Strategies of Aflatoxin Contamination
Althrough there is a fairly good understanding on the aflatoxin biosynthetic pathway and pathway cluster genes, many important questions still remain unanswered. Identification of all the genes responsible for aflatoxin formation and a better understanding of the regulatory mechanisms of aflatoxin formation of Aspergillus and the host crops will provide vital clues for devising strategies in solving aflatoxin contamination of food and feed.
Biological Control
Many natural isolates of A. flavus and A. parasiticus that do not produce aflatoxins. These non-aflatoxigenic strains have shown promise as a strategy to reduce preharvest aflatoxin contamination of crops (Dorner, 2005). Some of the non-aflatoxin-producing strains can out-compete the toxigenic strains in nature, and the strategy of competitive exclusion has been successfully practiced in cotton, peanut, and corn fields (Dorner, 2005). Cotty (1990) studied the competitiveness of seven non-toxigenic A. flavus strains in cotton in greenhouse experiments by co-inoculating them with representative toxigenic strains. Strain AF36 showed the highest survivability in greenhouse and field experiments and showed the largest reduction in aflatoxin levels. The EPA has approved AF36 as a biological control strain for field applications (Cotty, 1994). This strategy has also been shown to be effective in peanut and corn field studies (Dorner, 2005).
Host resistance through crop breeding and genetic engineering
Aspergillus flavus and A. parasiticus fungi can be found virtually everywhere in the world and are especially common in agricultural crop fields. Aflatoxin contamination tends to be greater when crops are subjected to insect damage and/or heat and drought stress. Various approaches have been suggested for genetic control of preharvest aflatoxin contamination including the development and use of crops with resistance to insects, resistance to plant stress (especially for tolerance to drought and high temperatures). Several sources of resistant germplasm were identified and released for crop genetic improvement in corn (McMillian et al., 1993; Williams and Windham, 2001; Naidoo et al., 2002; Guo et al. 2007a) and in peanuts (Cole et al., 1995; Holbrook et al., 2000; Guo et al., 2006; Holbrook et al., 2008). Crop resistance to aflatoxin contamination may be achieved by the three strategies: a) resistance to fungal invasion, b) inhibition of aflatoxin formation, and 3) resistance to insects and abiotic stress such as drought. Genetic engineering has been used in the last decade to improve crop resistance to diseases and insects. Examples are the development of Bt (Bacillus thuringiensis) corn (Ostlie et al., 1997; Singsit et al., 1996) and Bt peanut (Ozias-Akins et al., 2002). Transgenic Bt corn and Bt peanut have shown reduced levels of aflatoxin contamination (Windham et al., 1999; Dowd, 2001; Weissinger et al., 2002).
Functional genomics as a tool in reducing aflatoxin contamination
Peanut genomics research can provide new tools to study plant-microbe interactions and enhance crop genetic characteristics. However, genomics research in peanut is behind many other crops due to the shortage of essential genome infrastructure, tools, and resources. Peanut genomic work has involved the development of markers, maps, and the classification of species. However, improvement of methods for gene and genome sequencing and analysis of gene expression promise to allow rapid progress for identification and understanding of gene function related to phenotype. DNA sequencing is one of the most-important sources of genomic informaiton. The peanut genome (2,800 Mb/1C) is large in comparison to many model plants such as Arabidopsis (128 Mb), rice (420 Mb), and Medicago truncatula Gaertn. (500 Mb); and is larger than soybean Glycine max (L.) Merr. (1,100 Mb) and maize Zea mays L. (2,500 Mb) (Arumuganathan and Earle, 1991). The large genome size makes it unlikely that the peanut genome will be completely sequenced in the near future.
Sequencing of large numbers of Expressed Sequence Tags (ESTs) can deliver substantial amounts of genetic information on protein-coding sequences for comparative and functional genomic studies. ESTs provide putative genes, markers, and microarray resources to study peanut-Aspergillus interactions and to devise more effective breeding strategies to mitigate aflatoxin contamination. As of 5 October, 2007, a large number of ESTs of the top six plant species including Arapidopsis (1,276,692), rice (1,214,083), maize (1,159,264), wheat (1,050,932), oilseed rape (Brassica napus) (567,177) and barley (437,713) had been deposited to the GenBank database (dbEST release 100507, http://www.ncbi.nlm.nih.gov/dbEST/dbEST_summary.html). However, only 41,372 Arachis ESTs were deposited as of 5 October, 2007, in GenBank, including 6,264 from the wild species A. stenosperma (Krapov. and W.C. Gregory.). These ESTs were from different tissues, some of which had been subjected to different abiotic and biotic stresses (Luo et al., 2005a; Yan et al., 2005; Proite et al., 2007). Two submissions were 7454 and 2184 sequences from seed libraries of the Chinese accessions Luhua 14 and Shanyou 523, respectively. The majority represented the 21,777 ESTs (accession numbers ES702769 to ES724546) from developing seeds (Guo et al., 2008a) of the U.S. runner type cultivar Tifrunner (Holbrook and Culbreath, 2007) and the Spanish breeding line GT-C20 (Guo et al., 2008b). The seed and leaf EST originated from 10 non-normalized peanut cDNA libraries (Guo et al., 2007b; Guo et al., 2008a), resulting in over 13,824 unigene ESTs, which will be used for the production of a long-oligo microarray for peanut. This sequence data has been made available to the community in order to develop genomic tools and resources for deciphering the biological function of genes in the peanut genome, including a) construction of peanut 70-mer oligo microarray, and b) development of markers/genes associated with resistance to biotic and abiotic stress. A total of 8402 oligos of 60-80bp in length have been designed for the oligo microarray (Guo et al., 2007b).
Peanut ESTs developed from developing seeds and leaf tissues
Six different cDNA libraries were constructed from developing peanut seeds at three reproduction stages (R5, R6 and R7) from resistant and susceptible peanut genotypes. Tifrunner is susceptible to Aspergillus infection with higher aflatoxin contamination and resistant to Tomato Spotted Wilt Virus (TSWV) and GT-C20 is resistant to Aspergillus with reduced aflatoxin contamination and susceptible to TSWV. The developing peanut seed tissues were challenged by A. parasiticus and drought stress in the field. A total of 24,192 randomly selected cDNA clones from six libraries were sequenced. After removing vector sequences and quality trimming, 21,777 high-quality EST sequences were generated. Sequence clustering and assembling resulted in 8,689 unique EST sequences with 1,741 tentative consensus EST sequences (TCs) and 6,948 singleton ESTs. Functional classification was performed according to MIPS functional catalogue criteria. The unique EST sequences were divided into 22 categories. A similarity search against the non-redundant protein database available from NCBI (National Center for Biotechnology Information) indicated that 84.78% of the ESTs showed significant similarity to known proteins, of which 165 genes had been previously reported in peanuts. Furthermore, a comparison of these EST sequences with other plant sequences in The Institute for Genomic Research (TIGR) gene indices libraries showed that the percentage of peanut ESTs that matched to A. thaliana, maize (Zea mays L.), M. truncatula, rapeseed (Brassica napus L.), rice, soybean and wheat (Triticum aestivum (L.) Thell.) ESTs ranged from 33.84% to 79.46% with the sequence identity ≥ 80%. As expected, the peanut ESTs were more closely related to legume species than to cereal crops, and more homologous to dicot than to monocot plant species.
Two cDNA libraries were derived from peanut leaf tissues of the same peanut genotypes, Tifrunner and GT-C20, and 17,376 randomly picked clones were sequenced (Guo et al., unpublished data). After removing vector sequences and discarding low quality sequences, a total of 14,432 high-quality EST sequences were generated. Sequence clustering and assembling resulted in 6,888 unique EST sequences with 1,703 tentative consensus EST sequences (TCs) and 5,185 singleton ESTs. Functional annotations were performed on the basis of MIPS functional catalogue criteria and Gene Ontology (GO). According to the MIPS, about 82.9% (5,717) of the unique sequences have no known putative functions; and the matched sequences can be sorted into 15 categories, including 1.2% with defense-related genes. The unique EST sequences from GT-C20 and Tifrunner were analyzed for their potential use for developing simple sequence repeat (SSR) markers. Among 3,976 GT-C20 and 2,912 Tifrunner unique sequences, 593 and 263 EST-SSRs were developed, respectively. The frequency of SSR-containing ESTs was 14.2% in GT-C20 and 8.41% in Tifrunner. On average, one SSR was found every 4.52 kb in GT-C20 and one every 5.89 kb in Tifrunner. Approximate 56.54% of the EST-derived SSRs were di-nucleotide, 35.59% were tri-nucleotide, and the remaining 7.87% was comprised of tetra-, penta- and hexa-nucleotide repeat motifs. Among the identified EST-SSRs, the two most abundant sequence motif types were AG and AT recovered both in GT-C20 and Tifrunner.
EST sequencing, the random sequencing of clones from cDNA libraries, is the most efficient approach for gaining an initial picture of genes in the peanut genome. Functional classification provides useful information about the different metabolic and regulatory pathways that may be associated with environmental stresses. The developed ESTs can be used to discover novel sequences or genes, to identify resistance-related genes and to detect the differences among alleles or markers between these resistant and susceptible peanut genotypes. Studies with ESTs have given rise to a new resource for the development of microsatellites in A. hypogaea. SSR-containing EST sequences ( He et al., 2005; Khanal et al., 2008) have considerable potential for comparative mapping. Additionally, this large collection of cultivated peanut EST sequences will make it possible to construct microarrays for gene expression studies and for further characterization of host resistance mechanisms.
Analysis of Gene Expression
Macroarray (nylon-based) and microarray (glass slide-based) screening methods allow the simultaneous determination of expression levels of thousands of genes, making it possible to obtain a global view of the transcriptional state in a cell or tissue, and to associate genes with functions or specific physiological conditions. Recently, microarray technology has been demonstrated in a small scale pilot studies (Luo et al., 2005b; Luo et al., 2005c). EST-derived cDNA microarrays of 400 unigenes were probed under different conditions. Luo et al. (2005c) identified 25 ESTs that were potentially associated with drought stress or that responded to A. parasiticus challenge, and further study is warranted. Likewise, 56 up-regulated transcripts were identified and confirmed by real-time PCR upon infection with Cercosporidium personatum (Berk. et Curt.) Deighton. A long oligonucleotide microarray consisting of more gene-elements is under development (Guo et al., 2007b). Luo et al. (2005c) reported differentially expressed genes using a cDNA microarray containing 768 spots of 384 cDNA unigenes. They compared two treatments (A. parasiticus infection and drought stress) in order to better understand the relationship of drought stress and aflatoxin contamination in peanut. This enabled them to screen for genes differentially expressed in different environments. The expression of some genes selected from the microarray analyses have been validated by using real-time PCR. Further evaluation for individual genes will be needed in more genotypes to confirm the possible association of the gene expression with the resistant phenotype.
Aspergillus flavus functional genomics
ESTs, whole genome sequencing, and microarray technologies are robust genomic tools that provide high throughput capabilities (Kim et al., 2003; Yu et al., 2004d). These should be available tools for identification of genes involved in aflatoxin production and for studying the regulatory mechanisms for their expression. An EST project has been completed using the wild type strain NRRL 3357 (ATCC# 20026). More than 26,110 cDNA clones from a normalized cDNA expression library were sequenced at TIGR. After assembly and annotation, 7,218 unique sequences were identified (Yu et al., 2004d). These EST sequences have been released to the public at the NCBI GenBank Database. The A. flavus Gene Index was constructed at TIGR (http://www.tigr.org) which is currently maintained and curated by the Dana Farber Cancer Institute (http://compbio.dfci.harvard.edu/tgi). From this EST database, an additional four new transcripts (hypB, hypC, hypD, and hypE) were identified. Several categories of genes were identified that could potentially be involved, directly or indirectly, in aflatoxin production, such as in global regulation, signal transduction, pathogenicity, virulence, and fungal development. A genomic DNA amplicon microarray consisting of 5002 gene-elements was also constructed at TIGR. Profiling of genes involved in aflatoxin formation using microarrays performed at the Southern Regional Research Center of USDA-ARS and at TIGR identified hundreds of genes that are significantly up or down regulated (Kim et al., 2003; OBrian et al., 2003; Price et al., 2006). Further study on their functional involvement in aflatoxin formation is underway.
Sequencing of the A. flavus whole genome has been completed. Primary assembly indicated that the A. flavus genome consists of eight chromosomes and the genome size is about 36.3 Mb. Annotation of the A. flavus genome sequence data, with the help of A. flavus EST database, demonstrated that there are over 12,000 functional genes in the A. flavus genome. Genes responsible for the biosynthesis of secondary metabolites such as aflatoxins are those encoding polyketide synthases, non-ribosomal peptide synthethases, cytochrome P450 monooxygenases, fatty acid synthases, carboxylases, dehydrogenases, reductases, oxidases, oxidoreductases, epoxide hydrolases, oxygenases, and methyltransferases (Yu et al., 2004c). Primary annotation revealed that there exist more than two dozen PKSs, many NRPS, and numerous cytochrome P450 enzymes in the A. flavus genome. A whole genome microarray has been constructed at TIGR. Comparative genome hybridization (CGH) and functional genomics studies using these genomic resources are and will be performed for genome analysis. A proposed microarray is under construction in TIGR to include 8,402 peanut genes and more than 12,000 A. flavus genes and will be made available to collegeaues to study crop-pathogen interaction and for identifying genes involved in both fungal invasion and crop resistance.
Conclusions
To reduce or eliminate preharvest and postharvest aflatoxin contamination in crops is a serious challenge facing scientists today. In this presentation, we have reviewed the discovery of the aflatoxin pathway genes, gene clusters, and genetics and genomics progress made in peanut and A. flavus. The availability of a peanut-Aspergillus microarray will give scientists a valuable tool for studying crop-pathogen interaction and for identifying genes involved in both fungal invasion and crop resistance. The rapid progress in genomic research of both host plants and fungal pathogens should lead to a better understanding of the mechanisms of aflatoxin formation, pathogenicity of the fungus, and crop-fungus interactions. Developing novel strategies to control aflatoxin contamination is the ultimate goal for scientist. Efforts are also being made to understand fungus biology and the mechanism of aflatoxin biosynthesis using genomic tools. This will contribute greatly to achieve the goal of devising novel strategies to eliminate preharvest aflatoxin contamination resulting in a safer food and feed supply.
Literature Cited
Arumuganathan K. and Earle E. D. 1991 Nuclear DNA content of some important plant species. Plant Mol. Biol. Rep 9 : 208 – 218 .
Bennett J. W. , Rubin P. L. , Lee L. S. , and Chen P. N. 1979 Influence of trace elements and nitrogen sources on versicolorin production by a mutant strain of Aspergillus parasiticus. Mycopathologia 69 : 161 – 166 .
Bennett J. W. 1981 Loss of norsolorinic acid and aflatoxin production by mutant of Aspergillus parasiticus. J. Gen. Microbiol 124 : 429 – 432 .
Bennett J. W. , Leong P. M. , Kruger S. J. , and Keyes D. 1986 Sclerotial and low aflatoxigenic morphological variants from haploid and diploid Aspergillus parasiticus. Experientia 42 : 848 – 851 .
Bennett J. W. and Papa K. E. 1988 The aflatoxigenic Aspergillus spp. Adv. Plant Pathol 6 : 265 – 279 .
Bhatnagar D.
,
Ehrlich K. C.
, and
Cleveland T. E.
1992
Oxidation-reduction reactions in biosynthesis of secondary metabolites.
255
–
285
In
Bhatnagar
,
et al
Mycotoxins in Ecological Systems
Marcel Dekker
New York
.
Blount W. P. 1961 Turkey X disease. J. Brit. Turkey Federation 9 : 55 .
Bressac B. , Kew M. , Wands J. , and Ozturk M. 1991 Selective G to T mutations of p53 gene in hepatocellular carcinoma from southern Africa. Nature 350 : 429 – 431 .
Brodhagen M. and Keller N. P. 2006 Signaling pathways connecting mycotoxin production and sporulation. Mol. Plant Pathol 7 : 285 – 301 .
Calvo A. M. , Wilson R. A. , Bok J. W. , and Keller N. P. 2002 Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev 66 : 447 – 459 .
CAST 1989 Mycotoxins, Economic and health Risks Ames, IA .
CAST 2003 Mycotoxins: Risk in Plant, Animal, and Human Systems Ames, IA .
CDC (Centers for Disease Control and Prevention) 2004 Outbreak of aflatoxin poisoning-eastern and central provinces, Kenya, January-July, 2004. MMWR Morb. Mortal. Weekly Rep 53 : 790 – 792 .
Chang P. K. , Skory C. D. , and Linz J. E. 1992 Cloning of a gene associated with aflatoxin B1 biosynthesis in Aspergillus parasiticus. Curr. Genet 21 : 231 – 233 .
Chang P. K. , Cary J. W. , Bhatnagar D. , Cleveland T. E. , Bennett J. W. , Linz J. E. , Woloshuk C. P. , and Payne G. A. 1993 Cloning of the Aspergillus parasiticus apa-2 gene associated with the regulation of aflatoxin biosynthesis. Appl. Environ. Microbiol 59 : 3273 – 3279 .
Chang P. K. , Bennett J. W. , and Cotty P. J. 2002 Association of aflatoxin biosynthesis and sclerotial development in Aspergillus parasiticus. Mycopathologia 153 : 41 – 48 .
Cole R. J. and Cole E. H. 1987 Handbook of Toxic Fungal Metabolites Academic Press New York .
Cole R. J.
,
Dorner J. W.
, and
Holbrook C. C.
1995
Advances in mycotoxin elimination and resistance.
456
–
474
In
Pattee H. E.
and
Stalker H. T.
Advance in Peanut Science
Amer. Peanut Res. Educ. Soc
Stillwater, OK
.
Cotty P. 1988 Aflatoxin and sclerotial production by Aspergillus flavus: Influence of pH. Phytopathology 78 : 1250 – 1253 .
Cotty P. J. 1990 Effect of atoxigenic strains of Aspergillus flavus on aflatoxin contamination of developing cottonseed. Plant Dis 74 : 233 – 235 .
Cotty P. J. 1994 Influence of field application of an atoxigenic strain of Aspergillus flavus on the populations of A. flavus infecting cotton bolls and on the aflatoxin content of cottonseed. Phytopathol 84 : 1270 – 1277 .
Cuero R. , Ouellet T. , Yu J. , and Mogongwa N. 2003 Metal ion enhancement of fungal growth, gene expression and aflatoxin synthesis in Aspergillus flavus: RT-PCR characterization. J. Appl. Microbiol 94 : 953 – 961 .
Dorner J. W.
2005
Biological control of aflatoxin crop contamination.
207
–
215
In
Abbas H.
Aflatoxin and Food Safety
CRC Press
Boca Raton, FL
.
Dowd P. F. 2001 Biotic and abiotic factors limiting efficacy of Bt corn in indirectly reducing mycotoxin levels in commercial fields. J. Econ. Entomol 94 : 1067 – 1074 .
Eaton D. L. and Groopman J. D. 1994 The Toxicology of Aflatoxins: Human Health, Veterinary and Agricultural Significance Academic Press New York .
Ehrlich K. C. , Chang P. K. , Yu J. , and Cotty P. J. 2004 Aflatoxin biosynthesis cluster gene cypA is required for G aflatoxin formation. Appl. Environ. Microbiol 70 : 6518 – 6524 .
Feng G. H. and Leonard T. J. 1998 Culture conditions control expression of the genes for aflatoxin and sterigmatocystin biosynthesis in Aspergillus parasiticus and A. nidulans. Appl. Environ. Microbiol 64 : 2275 – 2277 .
Forgacs J. and Carll W. T. 1962 Mycotoxicoses. Advan. Vet. Sci 7 : 273 – 382 .
Goto T. , Wicklow D. T. , and Ito Y. 1996 Aflatoxin and cyclopiazonic acid production by a sclerotium-producing Aspergillus tamarii strain. Appl. Environ. Microbiol 62 : 4036 – 4038 .
Guo B. Z. , Chen X. , Dang P. , Scully B. T. , Liang X. , Holbrook C. C. , Yu J. , and Culbreath A. K. 2008a Peanut gene expression profiling in developing seeds at different reproduction stages during Aspergillus parasiticus infection. BMC Developmental Biology 8 : 12 .
Guo B. Z.
,
Holbrook C. C.
,
Yu J.
,
Lee R. D.
, and
Lynch R. E.
2005a
Application of technology of gene expression in response to drought stress and elimination of preharvest aflatoxin contamination.
313
–
331
In
Abbas H. K.
Aflatoxin and Food Safety
CRC Press
Boca Raton, FL
.
Guo B. Z.
,
Liang X.
,
Chung S-Y.
,
Holbrook C. C.
, and
Maleki S. J.
2008b
Proteomic analysis of peanut seed storage proteins and genetic variation in a potential peanut allergen.
15
:
567
–
577
Protein and Peptide Letters
.
Guo B. Z.
,
Widstrom N. W.
,
Lee R. D.
,
Wilson D. M.
, and
Coy A. E.
2005b
Prevention of preharvest aflatoxin contamination: Integration of crop management and genetic in corn.
437
–
457
In
Abbas H.
Aflatoxin and Food Safety
CRC Press
Boca Raton, FL
.
Guo B. Z. , Xu G. , Cao Y. G. , Holbrook C. C. , and Lynch R. E. 2006 Identification and characterization of phospholipase D and its association with drought susceptibilities in peanut (Arachis hypogaea). Planta 223 : 512 – 520 .
Guo B. Z. , Widstrom N. W. , Lee R. D. , Coy A. E. , and Lynch R. E. 2007a Registration of maize germplasm GT601 (AM-1) and GT602 (AM-2). J. Plant Registrations 1 : 153 – 154 .
Guo B. Z.
,
Chen X.
,
Dang P.
,
Holbrook C.
,
Culbreath A.
,
Scully B.
,
Yu J.
,
Nierman W. C.
, and
Cleveland T. E.
2007b
Generation of expressed sequence tags (ESTs) from cultivated peanuts for gene discovery and marker development.
Proc. Annual Aflatoxin Workshop
Oct. 21-14, 2007,
Atlanta, GA
.
He G. , Meng R. , Gao H. , Guo B. , Gao G. , Newman M. , Pittman R. , and Prakash C. S. 2005 Simple sequence repeat markers for botanical varieties of cultivated peanut (Arachis hypogaea L.). Euphytica 142 : 131 – 136 .
Henry S. H. , Bosch F. X. , Troxell T. C. , and Bolger P. M. 1999 Policy forum: Public health. Reducing liver cancer–global control of aflatoxin. Science 286 : 2453 – 2454 .
Hicks J. K. , Yu J. H. , Keller N. P. , and Adams T. H. 1997 Aspergillus sporulation and mycotoxin production both require inactivation of the FadA G alpha protein-dependent signaling pathway. EMBO J 16 : 4916 – 4923 .
Holbrook C. C. and Culbreath A. K. 2007 Registration of ‘Tifrunner’ peanut. J. Plant Registrations 1 : 124 .
Holbrook C. C. , Guo B. Z. , Wilson D. M. , and Timper P. 2009 The U.S. breeding program to develop peanut with drought tolerance and reduced aflatoxin contamination. Peanut Sci 36 : 49 – 52 .
Holbrook C. C. , Kvien C. K. , Rucker K. S. , Wilson D. M. , Hook J. E. , and Matheron M. E. 2000 Preharvest aflatoxin contamination in drought tolerant and drought intolerant peanut genotypes. Peanut Sci 27 : 45 – 48 .
Hsu I. C. , Metcalf R. A. , Sun T. , Welsh J. A. , Wang N. J. , and Harris C. C. 1991 Mutational hotspot in the p53 gene in human hepatocellular carcinomas. Nature 350 : 427 – 428 .
Khanal S.
,
Tang S.
,
Beilinson V.
,
Miguel P. S.
,
Guo B.
,
Nielsen N.
,
Stalker T.
,
Pratt M-M. C.
,
Pratt L. H.
,
Johnson V. E.
,
Taylor C. A.
, and
Knapp S. J.
2008
ESTs supply a wealth of polymorphic SSRs for genomics and molecular breeding applications in cultivated peanut.
Internal Plant and Animal Genome XVI Conference, January 12-16, 2008,
San Diego, CA
.
Kim H. , Snesrud E. C. , Haas B. , Cheung F. , Town C. D. , and Quackenbush J. 2003 Gene expression analyses of Arabidopsis chromosome 2 using a genomic DNA amplicon microarray. Genome Res 13 : 327 – 340 .
Kim J. H. , Campbell B. C. , Yu J. , Mahoney N. , Chan K. L. , Molyneux R. J. , Bhatnagar D. , and Cleveland T. E. 2005 Examination of fungal stress response genes using Saccharomyces cerevisiae as a model system: Targeting genes affecting aflatoxin biosynthesis by Aspergillus flavus Link. Appl. Microbiol. Biotechnol 67 : 807 – 815 .
Kim J. H. , Campbell B. C. , Molyneux R. , Mahoney N. , Chan K. L. , Yu J. , Wilkinson J. R. , Cary J. , Bhatnagar D. , and Cleveland T. E. 2006 Gene targets for fungal and mycotoxin control. Mycotoxin Res 22 : 3 – 8 .
Krishnamachari K. A. , Bhat R. V. , Nagarajan V. , and Tilak T. B. 1975 Hepatitis due to aflatoxicosis: An outbreak of hepatitis in parts of western India. Lancet 1 : 1061 – 1063 .
Lewis L. , Onsongo M. , Njapau H. , Schurz-Rogers H. , Luber G. , Kieszak S. , Nyamongo J. , Backer L. , Dahiye A. M. , Misore A. , DeCock K. , and Rubin C. 2005 Aflatoxin contamination of commercial maize products during an outbreak of acute aflatoxicosis in eastern and central Kenya. Environ. Health Perspect 113 : 1763 – 1767 .
Luo M. , Dang P. , Guo B. Z. , He G. , Holbrook C. C. , Bausher M. G. , and Lee R. D. 2005a Generation of expressed sequence tags (ESTs) for gene discovery and marker development in cultivated peanut. Crop Sci 45 : 346 – 353 .
Luo M. , Dang P. , Holbrook C. C. , Bausher M. G. , Lee R. D. , Lynch R. E. , and Guo B. Z. 2005b Identification of transcripts involved in resistance responses to leaf spot disease caused by Cercosporidium personatum in peanut (Arachis hypogaea L.). Phytopathology 95 : 381 – 387 .
Luo M. , Liang X. , Dang P. , Holbrook C. C. , Bausher M. G. , Lee R. D. , and Guo B. Z. 2005c Microarray-based screening of differentially expressed genes in peanut in response to Aspergillus parasiticus infection and drought stress. Plant Sci 169 : 695 – 703 .
Maggio-Hall L. A. , Wilson R. A. , and Keller N. P. 2005 Fundamental contribution of beta-oxidation to polyketide mycotoxin production in planta. Mol. Plant Microbe. Interact 18 : 783 – 93 .
Mahoney N. and Molyneux R. J. 2004 Phytochemical inhibition of aflatoxigenicity in Aspergillus flavus by constituents of walnut (Juglans regia). J. Agric. Food Chem 52 : 1882 – 1889 .
McMillian W. W. , Widstrom N. W. , and Wilson D. M. 1993 Registration of GT-MAS:gk maize germplasm. Crop Sci 33 : 882 .
Minto R. E. and Townsend C. A. 1997 Enzymology and molecular biology of aflatoxin biosynthesis. Chem. Rev 97 : 2537 – 2555 .
Naidoo G. , Forbes A. M. , Paul C. , White D. G. , and Rocheford T. R. 2002 Resistance of Aspergillus ear rot and aflatoxin accumulation in maize F1 hybrids. Crop Sci 42 : 360 – 364 .
OBrian G. R. , Fakhoury A. M. , and Payne G. A. 2003 Identification of genes differentially expressed during aflatoxin biosynthesis in Aspergillus flavus and Aspergillus parasiticus. Fungal Genet. Biol 39 : 118 – 127 .
Ostlie K. R.
,
Hutchison W. D.
, and
Hellmich R. L.
1997
Bt Corn and European corn borer.
NCR Publ. 602,
Univ. Minn
St. Paul, MN
.
Ozias-Akins P.
,
Yang H.
,
Gill R.
,
Fan H.
, and
Lynch R. E.
2002
Reduction of aflatoxin contamination in peanut - a genetic engineering approach.
151
–
160
In
Rajasekaran K.
,
et al
Crop Biotechnology. Ameri. Chem. Soc
Oxford Press
Washington, DC
.
Payne G. A. and Hagler W. M. 1983 Effect of specific amino acids on growth and aflatoxin production by Aspergillus parasiticus and Aspergillus flavus in defined media. Appl. Environ. Microbiol 46 : 805 – 812 .
Payne G. A. , Nystrom G. J. , Bhatnagar D. , Cleveland T. E. , and Woloshuk C. P. 1993 Cloning of the afl-2 gene involved in aflatoxin biosynthesis from Aspergillus flavus. Appl. Environ. Microbiol 59 : 156 – 162 .
Payne G. A. and Brown M. P. 1998 Genetics and physiology of aflatoxin biosynthesis. Annu. Rev. Phytopathol 36 : 329 – 362 .
Price M. S. , Yu J. , Nierman W. C. , Kim H. S. , Pritchard B. , Jacobus C. A. , Bhatnagar D. , Cleveland T. E. , and Payne G. A. 2006 The aflatoxin pathway regulator AflR induces gene transcription inside and outside of the aflatoxin biosynthetic cluster. FEMS Microbiol. Lett 255 : 275 – 279 .
Prieto R. and Woloshuk C. P. 1997 ord1, an oxidoreductase gene responsible for conversion of O- methylsterigmatocystin to aflatoxin in Aspergillus flavus. Appl. Environ. Microbiol 63 : 1661 – 1666 .
Proite K. , Leal-Bertioli S. C. , Bertioli D. J. , Moretzsohn M. C. , da Silva F. R. , Martins N. F. , and Guimaraes P. M. 2007 ESTs from a wild Arachis species for gene discovery and marker development. BMC Plant Biol 7 : 7 .
Reib J. 1982 Development of Aspergillus parasiticus and formation of aflatoxin B1 under the influence of conidiogenesis affecting compounds. Arch. Microbiol 133 : 236 – 238 .
Reverberi M. , Fabbri A. A. , Zjalic S. , Ricelli A. , Punelli F. , and Fanelli C. 2005 Antioxidant enzymes stimulation in Aspergillus parasiticus by Lentinula edodes inhibits aflatoxin production. Appl. Microbiol. Biotechnol 69 : 207 – 215 .
Reverberi M. , Zjalic S. , Racelli A. , Fabbri A. A. , and Fanelli C. 2006 Oxidant/antioxidant balance in Aspergillus parasiticus affects aflatoxin biosynthesis. Mycotoxin Res 22 : 39 – 47 .
Robens J. F.
and
Cardwell K.
2005
The cost of mycotoxin management in the United States.
1
–
12
In
Abbas H. K.
Aflatoxin and Food Safety
CRC Press
Boca Raton, FL
.
Singsit C. , Adang M. J. , Lynch R. E. , Anderson W. F. , Wang A. , Cardineau G. , and Ozias-Akins P. 1996 Expression of a Bacillus thuringiensis cryIA8 gene in transgenic peanut plants and its efficacy against lesser cornstalk borer. Transgenic Res 5 : 1 – 8 .
Skory C. D. , Chang P. K. , and Linz J. E. 1993 Regulated expression of the nor-1 and ver-1 genes associated with aflatoxin biosynthesis. Appl. Environ. Microbiol 59 : 1642 – 1646 .
Sobolev V. S. , Guo B. Z. , Holbrook C. C. , and Lynch R. E. 2007 Interrelationship of phytoalexin production and disease resistance in selected peanut genotypes. J. Agric. Food Chem 55 : 2195 – 2200 .
Trail F. , Mahanti N. , and Linz J. 1995a Molecular biology of aflatoxin biosynthesis. Microbiology 141 : 755 – 765 .
Trail F. , Mahanti N. , Rarick M. , Mehigh R. , Liang S. H. , Zhou R. , and Linz J. E. 1995b Physical and transcriptional map of an aflatoxin gene cluster in Aspergillus parasiticus and functional disruption of a gene involved early in the aflatoxin pathway. Appl. Environ. Microbiol 61 : 2665 – 2673 .
Van Egmond H. P. 1989 Current situation on regulations for mycotoxins. Overview of tolerances and status of standard methods of sampling and analysis. Food Addit. Contam 6 : 139 – 188 .
Watanabe C. M. and Townsend C. A. 2002 Initial characterization of a type I fatty acid synthase and polyketide synthase multienzyme complex NorS in the biosynthesis of aflatoxin B1. Chem. Biol 9 : 981 – 988 .
Weissinger A. , Wu M. , Liu Y-S. , Ingram K. , Rajasekaran K. , and Cleveland T. E. 2002 Development of transgenic peanut with enhanced resistance against preharvest aflatoxin contamination. Mycopathologia 155 : 97 .
Wilkinson J. R. , Yu J. , Bland J. M. , Nierman W. C. , Bhatnagar D. , and Cleveland T. E. 2007 Amino acid supplementation reveals differential regulation of aflatoxin biosynthesis in Aspergillus flavus NRRL 3357 and Aspergillus parasiticus SRRC 143. Appl. Microbiol. Biotechnol 74 : 1308 – 1319 .
Williams W. P. and Windham G. L. 2001 Registration of maize germplasm line Mp715. Crop Sci 41 : 1374 – 1375 .
Windham G. L. , Williams W. P. , and Davis F. M. 1999 Effects of the southwestern corn borer on Aspergillus flavus kernel infection and aflatoxin accumulation in maize hybrids. Plant Dis 83 : 535 – 540 .
Yabe K. , Ando Y. , and Hamasaki T. 1988 Biosynthetic relationship among aflatoxins B1, B2, G1, and G2. Appl. Environ. Microbiol 54 : 2101 – 2106 .
Yabe K. , Chihaya N. , Hamamatsu S. , Sakuno E. , Hamasaki T. , Nakajima H. , and Bennett J. W. 2003 Enzymatic conversion of averufin to hydroxyversicolorone and elucidation of a novel metabolic grid involved in aflatoxin biosynthesis. Appl. Environ. Microbiol 69 : 66 – 73 .
Yan Y. S. , Lin X. D. , Zhang Y. S. , Wang L. , Wu K. Q. , and Huang S. Z. 2005 Isolation of peanut genes encoding arachins and conglutins by expressed sequence tags. Plant Sci 169 : 439 – 445 .
Yu J. , Chang P. K. , Ehrlich K. C. , Cary J. W. , Montalbano B. , Dyer J. M. , Bhatnagar D. , and Cleveland T. E. 1998 Characterization of the critical amino acids of an Aspergillus parasiticus cytochrome P-450 monooxygenase encoded by ordA that is involved in the biosynthesis of aflatoxins B1, G1, B2, and G2. Appl. Environ. Microbiol 64 : 4834 – 4841 .
Yu J.
2004
Genetics and biochemistry of mycotoxin synthesis.
343
–
361
In
Arora D. K.
Fungal Biotechnology in Agricultural, Food, and Environmental Applications
Marcel Dekker
New York
.
Yu J. , Bhatnagar D. , and Cleveland T. E. 2004a Completed sequence of aflatoxin pathway gene cluster in Aspergillus parasiticus. FEBS Lett 564 : 126 – 130 .
Yu J.
,
Bhatnagar D.
, and
Cleveland T. E.
2004b
Genetics and biochemistry of aflatoxin formation and genomics approach for eliminating aflatoxin contamination.
224
–
242
In
Romeo J. T.
Recent Advances in Phytochemistry
Elsevier Publ. Co
New York
.
Yu J. , Chang P. K. , Ehrlich K. C. , Cary J. W. , Bhatnagar D. , Cleveland T. E. , Payne G. A. , Linz J. E. , Woloshuk C. P. , and Bennett J. W. 2004c Clustered pathway genes in aflatoxin biosynthesis. Appl. Environ. Microbiol 70 : 1253 – 1262 .
Yu J. , Whitelaw C. A. , Nierman W. C. , Bhatnagar D. , and Cleveland T. E. 2004d Aspergillus flavus expressed sequence tags for identification of genes with putative roles in aflatoxin contamination of crops. FEMS Microbiol. Lett 237 : 333 – 340 .
Yu J. , Cleveland T. E. , Nierman W. C. , and Bennett J. W. 2005 Aspergillus flavus genomics: Gateway to human and animal health, food safety, and crop resistance to diseases. Rev. Iberoam Micol 22 : 194 – 202 .
Yu J. , Cleveland T. E. , Wilkinson J. R. , Campbell B. C. , Kim J. H. , Kim H. S. , Bhatnagar D. , Payne G. A. , and Nierman W. C. 2006 Aspergillus flavus expressed sequence tags and microarray as tools in understanding aflatoxin biosynthesis. Mycotoxin Res 22 : 16 – 21 .
Yu J. H. 2006 Heterotrimeric G protein signaling and RGSs in Aspergillus nidulans. J. Microbiol 44 : 145 – 154 .
Notes
Author Affiliations
1 USDA/ARS, Crop Protection and Management Research Unit, Tifton, GA 31793.
2 USDA/ARS, Southern Regional Research Center, New Orleans, LA 70124.
3 USDA-ARS, Crop Genetics and Breeding Research Unit, Tifton, GA 31793.
4 J. Craig Venter Institute, Rockville, MD 20850.
5 Department of Biochemistry and Molecular Biology, The George Washington University School of Medicine, Washington, DC 20037, USA.
*Corresponding author (e-mail: Baozhu.Guo@ars.usda.gov)